Ce studiază biologia moleculară. Biologie moleculară - Biologie moleculară. Baza moleculară a eredității
Biologie moleculara a cunoscut o perioadă de dezvoltare rapidă a propriilor metode de cercetare, care acum diferă de biochimie. Acestea includ, în special, metode de inginerie genetică, clonare, expresie artificială și knockout genetic. Deoarece ADN-ul este purtătorul material al informațiilor genetice, biologia moleculară a devenit mult mai aproape de genetică, iar genetica moleculară s-a format la joncțiune, care este atât o secțiune a geneticii, cât și a biologiei moleculare. La fel cum biologia moleculară folosește pe scară largă virușii ca instrument de cercetare, virologia folosește metodele biologiei moleculare pentru a-și rezolva problemele. Pentru analiză informatii genetice este implicată tehnologia informatică, în legătură cu care au apărut noi direcții genetica moleculara care sunt uneori considerate discipline distincte: bioinformatica, genomica și proteomica.
Istoria dezvoltării
Această descoperire fundamentală a fost pregătită printr-o fază lungă de cercetare în genetica și biochimia virusurilor și bacteriilor.
În 1928, Frederick Griffith a arătat pentru prima dată că un extract de bacterii patogene ucise de căldură ar putea transfera trăsătura de patogenitate către bacteriile benigne. Studiul transformării bacteriene a condus în continuare la purificarea agentului bolii, care, contrar așteptărilor, s-a dovedit a fi nu o proteină, ci un acid nucleic. Acidul nucleic în sine nu este periculos, purtând doar genele care determină patogenitatea și alte proprietăți ale microorganismului.
În anii 50 ai secolului XX, s-a demonstrat că bacteriile au un proces sexual primitiv, sunt capabile să schimbe ADN extracromozomial, plasmide. Descoperirea plasmidelor, precum și transformările, au stat la baza tehnologiei plasmidelor comune în biologia moleculară. O altă descoperire importantă pentru metodologie a fost descoperirea la începutul secolului al XX-lea a virusurilor bacteriene, bacteriofage. Fagii pot transfera, de asemenea, material genetic de la unul celula bacteriana altcuiva. Infecția bacteriilor de către fagi duce la o modificare a compoziției ARN bacterian. Dacă, fără fagi, compoziția ARN-ului este similară cu compoziția ADN-ului bacterian, atunci după infecție, ARN-ul devine mai asemănător cu ADN-ul bacteriofag. Astfel, s-a constatat că structura ARN-ului este determinată de structura ADN-ului. La rândul său, rata sintezei proteinelor în celule depinde de cantitatea de complexe ARN-proteină. Așa a fost formulat Dogma centrală a biologiei moleculare: ADN ↔ ARN → proteină.
Dezvoltarea ulterioară a biologiei moleculare a fost însoțită atât de dezvoltarea metodologiei sale, în special, de inventarea unei metode de determinare a secvenței de nucleotide a ADN-ului (W. Gilbert și F. Sanger, Premiul Nobel pentru Chimie 1980), cât și de noi descoperiri în domeniul cercetării în structura și funcționarea genelor (vezi Istoria Geneticii). LA începutul lui XXI secolului, s-au obținut date despre structura primară a întregului ADN uman și a unui număr de alte organisme, cele mai importante pentru medicină, agricultură și cercetarea științifică, ceea ce a dus la apariția mai multor domenii noi în biologie: genomica, bioinformatica etc.
Vezi si
- biologie moleculară (jurnal)
- Transcriptomica
- Paleontologie moleculară
- EMBO - Organizația Europeană biologi moleculari
Literatură
- Cântărețul M., Berg P. Gene și genoame. - Moscova, 1998.
- Stent G., Kalindar R. Genetica moleculara. - Moscova, 1981.
- Sambrook J., Fritsch E.F., Maniatis T. Clonarea moleculară. - 1989.
- Patrushev L.I. Exprimarea genelor. - M.: Nauka, 2000. - 000 p., ill. ISBN 5-02-001890-2
Legături
Fundația Wikimedia. 2010 .
- Districtul Ardatovsky din regiunea Nijni Novgorod
- Districtul Arzamas din regiunea Nijni Novgorod
Vedeți ce este „Biologie moleculară” în alte dicționare:
BIOLOGIE MOLECULARA- studiază elementele de bază. proprietăţile şi manifestările vieţii la nivel molecular. Cele mai importante direcții în M. b. sunt studii structurale organizare functionala aparatul genetic al celulelor și mecanismul de implementare a informațiilor ereditare ... ... Dicționar enciclopedic biologic
BIOLOGIE MOLECULARA- explorează proprietățile și manifestările de bază ale vieții la nivel molecular. Afla cum și în ce măsură creșterea și dezvoltarea organismelor, stocarea și transmiterea informațiilor ereditare, conversia energiei în celulele vii și alte fenomene se datorează ... Dicţionar enciclopedic mare
BIOLOGIE MOLECULARA Enciclopedia modernă
BIOLOGIE MOLECULARA- BIOLOGIA MOLECULARA, studiul biologic al structurii si functiei MOLECULELOR care alcatuiesc organismele vii. Principalele domenii de studiu sunt fizice și Proprietăți chimice proteine și ACIDI NUCLEICI precum ADN-ul. Vezi si… … Dicționar enciclopedic științific și tehnic
biologie moleculara- o secțiune de biol., care explorează proprietățile și manifestările de bază ale vieții la nivel molecular. Afla cum și în ce măsură creșterea și dezvoltarea organismelor, stocarea și transmiterea informațiilor ereditare, conversia energiei în celulele vii și ... ... Dicţionar de microbiologie
biologie moleculara- — Subiecte de biotehnologie EN biologie moleculară … Manualul Traducătorului Tehnic
Biologie moleculara- BIOLOGIA MOLECULARA, exploreaza proprietatile si manifestarile de baza ale vietii la nivel molecular. Afla cum și în ce măsură creșterea și dezvoltarea organismelor, stocarea și transmiterea informațiilor ereditare, conversia energiei în celulele vii și ... ... Dicţionar Enciclopedic Ilustrat
Biologie moleculara- o știință care își pune ca sarcină cunoașterea naturii fenomenelor vieții prin studierea obiectelor și sistemelor biologice la un nivel apropiat de nivelul molecular, și în unele cazuri atingând această limită. Scopul final al acestui lucru este…… Marea Enciclopedie Sovietică
BIOLOGIE MOLECULARA- studiază fenomenele vieții la nivelul macromoleculelor (cap. arr. proteine și acizi nucleici) în structurile libere de celule (ribozomi etc.), în virusuri, dar și în celule. scopul lui M. stabilirea rolului și mecanismului de funcționare a acestor macromolecule pe baza ...... Enciclopedia chimică
biologie moleculara- explorează proprietățile și manifestările de bază ale vieții la nivel molecular. Afla cum și în ce măsură creșterea și dezvoltarea organismelor, stocarea și transmiterea informațiilor ereditare, conversia energiei în celulele vii și alte fenomene ... ... Dicţionar enciclopedic
Cărți
- Biologia moleculară a celulei. Cartea cu probleme, J. Wilson, T. Hunt. Cartea autorilor americani este o anexă la cea de-a 2-a ediție a manualului `Molecular Biology of the Cell` de B. Alberts, D. Bray, J. Lewis și alții. Conține întrebări și sarcini, al căror scop este aprofundarea ...
Se poate spune că biologia moleculară studiază manifestările vieții pe structuri sau sisteme neînsuflețite cu semne elementare de activitate vitală (care pot fi macromolecule biologice individuale, complexele sau organitele acestora), studiind modul în care procesele cheie care caracterizează materia vie sunt realizate prin interacțiuni și transformări chimice.
Separarea biologiei moleculare de biochimie într-un domeniu independent de știință este dictată de faptul că sarcina sa principală este de a studia structura și proprietățile macromoleculelor biologice implicate în diferite procese, pentru a elucida mecanismele interacțiunii lor. Biochimia, pe de altă parte, se ocupă cu studiul proceselor reale ale activității vitale, modelele cursului lor într-un organism viu și transformările moleculelor care însoțesc aceste procese. În cele din urmă, biologia moleculară încearcă să răspundă la întrebarea de ce are loc acest sau acel proces, în timp ce biochimia răspunde la întrebările unde și cum, din punctul de vedere al chimiei, are loc procesul în cauză.
Poveste
Biologia moleculară ca domeniu separat al biochimiei a început să prindă contur în anii 1930. Atunci, pentru o înțelegere mai profundă a fenomenului vieții, a apărut necesitatea unor studii țintite la nivel molecular ale proceselor de stocare și transmitere a informațiilor ereditare în organismele vii. Apoi, sarcina biologiei moleculare a fost determinată în studiul structurii, proprietăților și interacțiunii acizi nucleici si proteine. Termenul de „biologie moleculară” a fost folosit pentru prima dată de omul de știință englez William Astbury în contextul cercetărilor legate de elucidarea relației dintre structura moleculară și proprietățile fizice și biologice ale proteinelor fibrilare, cum ar fi colagenul, fibrina din sânge sau proteinele contractile musculare.
În primele zile ale biologiei moleculare, ARN-ul era considerat o componentă a plantelor și ciupercilor, în timp ce ADN-ul era văzut ca o componentă tipică a celulelor animale. Primul cercetător care a demonstrat că ADN-ul se găsește în plante a fost Andrey Nikolaevich Belozersky, care a izolat ADN-ul de mazăre în 1935. Această descoperire a stabilit faptul că ADN-ul este un acid nucleic universal prezent în celulele vegetale și animale.
O realizare majoră a fost stabilirea de către George Beadle și Edward Tatum a unei relații cauzale directe între gene și proteine. În experimentele lor, au expus celulele neurosporilor ( Neurosporacrassa) Expunerea la raze X care a provocat mutații. Rezultatele obținute au arătat că acest lucru a dus la o modificare a proprietăților unor enzime specifice.
În 1940, Albert Claude a izolat granule care conțin ARN citoplasmatic din citoplasma celulelor animale, care erau mai mici decât mitocondriile. Le-a numit microzomi. Ulterior, în studiul structurii și proprietăților particulelor izolate a fost stabilit rolul fundamental al acestora în procesul de biosinteză a proteinelor. În 1958, la primul simpozion dedicat acestor particule, s-a decis denumirea acestor particule ribozomi.
Un alt pas important în dezvoltarea biologiei moleculare au fost datele publicate ale experimentului lui Oswald Avery, Colin MacLeod și MacLean McCarthy în 1944, care au arătat că ADN-ul este cauza transformării bacteriene. Aceasta a fost prima dovadă experimentală a rolului ADN-ului în transmiterea informațiilor ereditare, dezmințind ideea anterioară a naturii proteice a genelor.
La începutul anilor 1950, Frederick Sanger a arătat că un lanț proteic este o secvență unică de reziduuri de aminoacizi. La sfârșitul anilor 1950, Max Perutz și John Kendrew au descifrat structura spațială a primelor proteine. Deja în anul 2000, erau cunoscute sute de mii de secvențe naturale de aminoacizi și mii de structuri spațiale ale proteinelor.
Aproximativ în același timp, cercetările lui Erwin Chargaff i-au permis să formuleze reguli care descriu raportul bazelor azotate din ADN (regulile spun că, indiferent de diferențele dintre specii în ADN, cantitatea de guanină este egală cu cantitatea de citozină, iar cantitatea de adenină este egală cu cantitatea de emină), ceea ce mai târziu a contribuit la realizarea celei mai mari descoperiri și a uneia dintre biologia moleculară. cele mai mari descopeririîn biologie în general.
Acest eveniment a avut loc în 1953 când James Watson și Francis Crick, bazat pe lucrarea lui Rosalind Franklin și Maurice Wilkins la Analiza difracției cu raze X ADN-ul, a stabilit structura dublu catenară a moleculei de ADN. Această descoperire a făcut posibil să se răspundă la întrebarea fundamentală despre capacitatea purtătorului de informații ereditare de a se auto-reproduce și de a înțelege mecanismul de transmitere a unor astfel de informații. Aceiași oameni de știință au formulat principiul complementarității bazelor azotate, care este de o importanță cheie pentru înțelegerea mecanismului de formare a structurilor supramoleculare. Acest principiu, care este folosit acum pentru a descrie toate complexele moleculare, face posibilă descrierea și prezicerea condițiilor de apariție a interacțiunilor intermoleculare slabe (nevalente), care determină posibilitatea formării de secundare, terțiare etc. structuri ale macromoleculelor, autoasamblarea supramoleculare sisteme biologice care determină o varietate atât de mare de structuri moleculare și seturile lor funcționale. Apoi, în 1953, a apărut Revista de Știință Jurnalul de Biologie Moleculară. A fost condus de John Kendrew, interese științifice care a fost studiul structurii proteinelor globulare (Premiul Nobel în 1962, împreună cu Max Perutz). O revistă similară în limba rusă numită Molecular Biology a fost fondată în URSS de V. A. Engelhardt în 1966.
În 1958, Francis Crick a formulat așa-numitul. Dogma centrală a biologiei moleculare: ideea ireversibilității fluxului de informații genetice de la ADN prin ARN la proteine, conform schemei ADN → ADN (replicare, crearea unei copii a ADN), ADN → ARN (transcripție, copiere a genelor), ARN → proteină (traducere, decodare a informațiilor despre structura proteinelor). Această dogmă a fost oarecum corectată în 1970, ținând cont de cunoștințele acumulate, deoarece fenomenul transcripției inverse a fost descoperit în mod independent de Howard Temin și David Baltimore: a fost descoperită o enzimă - transcriptaza inversă, care este responsabilă pentru implementarea transcripției inverse - formarea ADN-ului dublu catenar pe un virus monocatenar care are loc în ARN-ul cogenic. Trebuie remarcat faptul că stricta necesitate a fluxului de informații genetice de la acizi nucleici la proteine rămâne încă baza biologiei moleculare.
În 1957, Alexander Sergeevich Spirin, împreună cu Andrei Nikolaevich Belozersky, au arătat că, în ciuda diferențelor semnificative în compoziția de nucleotide a ADN-ului de la diferite organisme, compoziția ARN-ului total este similară. Pe baza acestor date, au ajuns la concluzia senzațională că ARN-ul total al unei celule nu poate acționa ca purtător de informații genetice de la ADN la proteine, deoarece nu îi corespunde în compoziția sa. În același timp, au observat că există o fracțiune minoră de ARN, care corespunde pe deplin în compoziția sa de nucleotide cu ADN-ul și care poate fi un adevărat purtător de informații genetice de la ADN la proteine. Drept urmare, ei au prezis existența unor molecule de ARN relativ mici, care sunt analoge ca structură cu secțiunile individuale de ADN și acționează ca intermediari în transferul informațiilor genetice conținute în ADN către ribozom, unde moleculele de proteine sunt sintetizate folosind aceste informații. În 1961 (S. Brenner, F. Jacob, M. Meselson, pe de o parte, și F. Gros, François Jacob și Jacques Monod, au fost primii care au confirmat experimental existența unor astfel de molecule - ARN informațional (matrice). În același timp, au dezvoltat conceptul și modelul unității funcționale a ADN-ului - operonul, ceea ce a făcut posibilă explicarea exactă a modului în care se desfășoară expresia proteinelor în studiul prokaryos al mecanismului biogenic. sinteza si principiile organizarea structurală iar munca mașinilor moleculare - ribozomi - a făcut posibilă formularea unui postulat care descrie mișcarea informației genetice, numită dogma centrală a biologiei moleculare: ADN - ARNm - proteină.
În 1961 și în următorii câțiva ani, Heinrich Mattei și Marshall Nirenberg, apoi Har Korana și Robert Holley, au efectuat mai multe lucrări de descifrare a codului genetic, în urma cărora s-a stabilit o relație directă între structura ADN-ului și proteinele sintetizate și s-a determinat secvența de nucleotide care determină setul de aminoacizi din proteină. S-au obţinut şi date despre universalitatea codului genetic. Descoperirile au primit Premiul Nobel în 1968.
Pentru dezvoltare ideile contemporane Despre funcțiile ARN, descoperirea ARN-ului necodificator, făcută pe baza rezultatelor lucrării lui Alexander Sergeevich Spirin împreună cu Andrei Nikolaevich Belozersky în 1958, Charles Brenner cu coautorii și Saul Spiegelman în 1961, a fost decisivă. Acest tip de ARN constituie cea mai mare parte a ARN-ului celular. ARN-urile ribozomale sunt în primul rând necodante.
Metodele de cultivare și hibridizare a celulelor animale au primit o dezvoltare serioasă. În 1963, François Jacob și Sydney Brenner au formulat ideea unui replicon, o secvență de gene care se reproduc în mod inerent, care explică aspecte importante ale reglării replicării genelor.
În 1967, în laboratorul lui A. S. Spirin, s-a demonstrat pentru prima dată că forma ARN-ului pliat compact determină morfologia particulei ribozomale.
În 1968, a fost făcută o descoperire fundamentală semnificativă. Okazaki, după ce a descoperit fragmente de ADN ale lanțului întârziat în studiul procesului de replicare, a numit fragmente Okazaki după ea, a clarificat mecanismul de replicare a ADN-ului.
În 1970, Howard Temin și David Baltimore au făcut în mod independent o descoperire semnificativă: a fost descoperită o enzimă - transcriptaza inversă, care este responsabilă pentru implementarea transcripției inverse - formarea ADN-ului dublu catenar pe un șablon ARN monocatenar, care apare în virusurile oncogene care conțin ARN.
încă una realizare importantă biologia moleculară a fost explicația mecanismului mutațiilor la nivel molecular. În urma unei serii de studii s-au stabilit principalele tipuri de mutații: dublări, inversiuni, deleții, translocații și transpoziții. Acest lucru a făcut posibilă luarea în considerare a schimbărilor evolutive din punctul de vedere al proceselor genice și a făcut posibilă dezvoltarea teoriei ceasurilor moleculare, care este folosită în filogenie.
Până la începutul anilor 1970, au fost formulate principiile de bază ale funcționării acizilor nucleici și proteinelor într-un organism viu. S-a constatat că proteinele și acizii nucleici din organism sunt sintetizați conform unui mecanism de matrice, molecula de matrice transportând informații criptate despre secvența de aminoacizi (într-o proteină) sau nucleotid (într-un acid nucleic). În timpul replicării (dublarea ADN-ului) sau transcripției (sintezei ARNm), ADN-ul servește ca un astfel de șablon, în timpul translației (sintezei proteinelor) sau transcripției inverse - ARNm.
Astfel, au fost create premisele teoretice pentru dezvoltarea domeniilor aplicate ale biologiei moleculare, în special, ingineria genetică. În 1972, Paul Berg, Herbert Bauer și Stanley Cohen au dezvoltat tehnologia de clonare moleculară. Apoi au fost primii care au obținut ADN recombinant in vitro. Aceste experimente remarcabile au pus bazele ingineriei genetice, iar anul acesta este considerat data nașterii acestei direcții științifice.
În 1977, Frederick Sanger și, în mod independent, Allan Maxum și Walter Gilbert au dezvoltat diferite metode pentru determinarea structura primara(secvențiere) ADN. Metoda Sanger, așa-numita metodă de terminare a lanțului, stă la baza metodei moderne de secvențiere. Principiul secvențierii se bazează pe utilizarea bazelor marcate care acționează ca terminatori într-o reacție de secvențiere ciclică. Această metodă a devenit larg răspândită datorită capacității de a efectua rapid analize.
1976 - Frederick. Sanger a descifrat secvența de nucleotide a ADN-ului fagului φΧ174 cu o lungime de 5375 de perechi de nucleotide.
1981 - Anemia cu celule falciforme devine prima boală genetică diagnosticată prin analiza ADN.
1982-1983 descoperirea funcției catalitice a ARN-ului în laboratoarele americane ale lui T. Check și S. Altman a schimbat ideile existente despre rolul exclusiv al proteinelor. Prin analogie cu proteinele catalitice - enzime, ARN-urile catalitice au fost numite ribozime.
1987 Keri Mullez a descoperit reacția în lanț a polimerazei, datorită căreia este posibilă creșterea artificială semnificativă a numărului de molecule de ADN în soluție pentru lucrări ulterioare. Astăzi este una dintre cele mai importante metode de biologie moleculară folosită în studiul bolilor ereditare și virale, în studiul genelor și în identificarea genetică și înrudirea etc.
În 1990, în același timp, trei grupuri de oameni de știință au publicat o metodă care a făcut posibilă obținerea rapidă a ARN-urilor sintetice active funcțional în laborator (ribozime sau molecule artificiale care interacționează cu diverși liganzi - aptameri). Această metodă se numește „evoluție in vitro”. Și la scurt timp după aceea, în 1991-1993 în laboratorul lui A.B. Chetverina a fost demonstrată experimental posibilitatea existenței, creșterii și amplificării moleculelor de ARN sub formă de colonii pe medii solide.
În 1998, aproape simultan, Craig Mello și Andrew Fire au descris mecanismul observat mai devreme în experimentele genetice cu bacterii și flori. interferența ARN, în care o moleculă mică de ARN dublu catenar duce la o suprimare specifică a expresiei genelor.
Descoperirea mecanismului interferenței ARN este de mare importanță. valoare practică pentru biologia moleculară modernă. Acest fenomen este utilizat pe scară largă în experimente științifice ca instrument de „oprire”, adică suprimarea expresiei genelor individuale. Un interes deosebit este faptul că această metodă permite suprimarea reversibilă (temporară) a activității genelor studiate. Se fac cercetări pentru aplicarea acestui fenomen în tratamentul bolilor virale, neoplazice, degenerative și metabolice. Trebuie remarcat faptul că, în 2002, au fost descoperiți mutanți ai virusurilor poliomielitei care pot evita interferența ARN, așa că este nevoie de o muncă mai minuțioasă pentru a dezvolta un adevărat metode eficiente tratament bazat pe acest fenomen.
În 1999-2001, mai multe grupuri de cercetători au determinat structura ribozomului bacterian cu o rezoluție de 5,5 până la 2,4 angstromi.
Articol
Realizările biologiei moleculare în cunoașterea naturii vii pot fi cu greu supraestimate. Un mare succes a fost obținut datorită conceptului de succes de cercetare: complex procese biologice sunt considerate din punctul de vedere al sistemelor moleculare individuale, ceea ce permite utilizarea unor metode fizico-chimice precise de cercetare. De asemenea, a atras multe minți mari din domenii conexe în acest domeniu al științei: chimie, fizică, citologie, virologie, care au avut și un efect benefic asupra amplorii și vitezei de dezvoltare a cunoștințelor științifice în acest domeniu. Descoperiri atât de importante precum determinarea structurii ADN-ului, descifrarea codului genetic și modificarea artificială direcționată a genomului au făcut posibilă înțelegerea mult mai profundă a specificului proceselor de dezvoltare ale organismelor și rezolvarea cu succes a numeroase probleme științifice, medicale și sociale importante și aplicate, care erau considerate de nerezolvat nu cu mult timp în urmă.
Subiectul de studiu al biologiei moleculare îl constituie în principal proteinele, acizii nucleici și complexele moleculare (mașini moleculare) bazate pe acestea și procesele la care participă.
Acizii nucleici sunt polimeri liniari constând din unități nucleotidice (compuși dintr-un zahăr cu cinci membri cu o grupare fosfat la al cincilea atom al ciclului și una dintre cele patru baze azotate) interconectate printr-o legătură esterică a grupărilor fosfat. Astfel, acidul nucleic este un polimer de pentoză fosfat cu baze azotate ca substituenți laterali. Compoziție chimică Lanțul de ARN diferă de ADN prin faptul că primul constă dintr-un ciclu de carbohidrați cu cinci membri, în timp ce al doilea constă dintr-un derivat de riboză dehidroxilat - deoxiriboză. În același timp, aceste molecule diferă dramatic în spațiu, deoarece ARN-ul este o moleculă flexibilă monocatenară, în timp ce ADN-ul este o moleculă dublu catenară.
Proteinele sunt polimeri liniari, care sunt lanțuri de alfa-aminoacizi legați între ele. legătură peptidică, de unde și al doilea lor nume - polipeptide. Compoziția proteinelor naturale include multe unități diferite de aminoacizi - la om până la 20 -, ceea ce determină o mare varietate de proprietăți funcționale ale acestor molecule. Acestea sau acele proteine participă la aproape fiecare proces din organism și îndeplinesc multe sarcini: joacă rolul de material de construcție celulară, asigură transportul de substanțe și ioni, catalizează reacții chimice, această listă este foarte lungă. Proteinele formează conformații moleculare stabile de diferite niveluri de organizare (structuri secundare și terțiare) și complexe moleculare, ceea ce le extinde și mai mult funcționalitatea. Aceste molecule pot avea o specificitate ridicată pentru îndeplinirea anumitor sarcini datorită formării unei structuri globulare spațiale complexe. O mare varietate de proteine asigură interesul constant al oamenilor de știință pentru acest tip de molecule.
Ideile moderne despre subiectul biologiei moleculare se bazează pe o generalizare prezentată pentru prima dată în 1958 de Francis Crick ca dogma centrală a biologiei moleculare. Esența sa a fost afirmația că informația genetică din organismele vii trece prin etape de implementare strict definite: copierea de la ADN la ADN la intrarea moștenirii, de la ADN la ARN și apoi de la ARN la proteină, iar tranziția inversă nu este fezabilă. Această afirmație a fost adevărată doar parțial, prin urmare, ulterior, dogma centrală a fost corectată cu privire la datele nou descoperite.
În prezent, există mai multe modalități de implementare a materialului genetic, reprezentând diferite secvențe pentru implementarea celor trei tipuri de existență a informației genetice: ADN, ARN și proteină. În nouă moduri posibile de realizare, se disting trei grupuri: acestea sunt trei transformări generale (generale), care se desfășoară în mod normal la majoritatea organismelor vii; trei transformari speciale (speciale), efectuate la unii virusi sau in conditii speciale de laborator; trei transformări necunoscute (necunoscute), a căror implementare este considerată imposibilă.
Transformările comune includ următoarele moduri de implementare a codului genetic: ADN→ADN (replicare), ADN→ARN (transcripție), ARN→proteină (traducere).
Pentru a efectua transferul trăsăturilor ereditare, părinții trebuie să transmită descendenților lor o moleculă de ADN cu drepturi depline. Procesul prin care o copie exactă a ADN-ului original poate fi sintetizată și, prin urmare, materialul genetic poate fi transferat, se numește replicare. Este realizat de proteine speciale care desfășoară molecula (își îndreptă secțiunea), se desfășoară dublu helixși folosind ADN polimeraza pentru a crea o copie exactă a moleculei de ADN originală.
Pentru a asigura viața unei celule, aceasta trebuie să se refere în mod constant la codul genetic încorporat în dubla helix ADN. Cu toate acestea, această moleculă este prea mare și stângace pentru a fi folosită ca sursă directă de material genetic pentru sinteza continuă a proteinelor. Prin urmare, în cursul implementării informațiilor încorporate în ADN, există o etapă intermediară: sinteza ARNm, care este o moleculă mică monocatenar complementară unui anumit segment de ADN care codifică o anumită proteină. Procesul de transcripție este asigurat de ARN polimerază și factori de transcripție. Molecula rezultată poate fi apoi livrată cu ușurință în partea celulei responsabilă de sinteza proteinelor - ribozomul.
După ce ARN intră în ribozom, începe etapa finală a realizării informației genetice. Ribozomul citește din ARNm cod genetic tripleti numiti codoni si sintetizeaza proteina corespunzatoare pe baza informatiilor primite.
În cursul transformărilor speciale, codul genetic se realizează după schema ARN → ARN (replicare), ARN → ADN (transcripție inversă), ADN → proteină (traducere directă). Replicarea acestui tip este realizată în mulți viruși, unde este realizată de enzima ARN polimeraza dependentă de ARN. Enzime similare se găsesc și în celulele eucariote, unde sunt asociate cu procesul de tăcere a ARN-ului. Transcripția inversă a fost găsită în retrovirusuri, unde este efectuată de enzima transcriptază inversă și, în unele cazuri, în celulele eucariote, de exemplu, în timpul sintezei telomerice. Transmiterea în direct se realizează numai în condiții artificiale într-un sistem izolat în afara celulei.
Oricare dintre cele trei posibile tranziții ale informațiilor genetice de la proteină la proteină, ARN sau ADN este considerată imposibilă. Cazul acțiunii prionilor asupra proteinelor, în urma căreia se formează un prion similar, ar putea fi atribuit condiționat tipului de realizare a informației genetice proteină → proteină. Cu toate acestea, în mod formal nu este așa, deoarece nu afectează secvența de aminoacizi din proteină.
Istoria apariției termenului de „dogma centrală” este curioasă. Deoarece cuvântul dogmă înseamnă în general o afirmație care nu este supusă îndoielii, iar cuvântul în sine are o conotație religioasă clară, alegându-l ca descriere fapt științific nu pe deplin legitime. Potrivit lui Francis Crick însuși, a fost greșeala lui. El a vrut să dea mai multă semnificație teoriei propuse, să o deosebească de fundalul altor teorii și ipoteze; de ce a decis să folosească acest cuvânt maiestuos, după părerea lui, neînțelegându-i adevăratul sens. Numele, însă, a rămas.
Biologia moleculară astăzi
Dezvoltarea rapidă a biologiei moleculare, interesul constant pentru realizările în acest domeniu din partea societății și importanța obiectivă a cercetării au dus la apariția un numar mare principalele centre de cercetare de biologie moleculară din întreaga lume. Printre cele mai mari, trebuie menționate următoarele: laboratorul de biologie moleculară din Cambridge, Institutul Regal din Londra - în Marea Britanie; institute de biologie moleculară din Paris, Marsilia și Strasbourg, Institutul Pasteur - în Franța; departamentele de biologie moleculară de la Universitatea Harvard și Institutul de Tehnologie din Massachusetts, Universitatea din Berkeley, Institutul de Tehnologie din California, Universitatea Rockefeller, Institutul de Sănătate Publică din Bethesda - în SUA; institutele Max Planck, universitățile din Göttingen și München, Institutul Central de Biologie Moleculară din Berlin, institutele din Jena și Halle - din Germania; Institutul Karolinska din Stockholm, Suedia.
În Rusia, centrele de frunte în acest domeniu sunt Institutul de Biologie Moleculară. Institutul de Genetică Moleculară RAS, Institutul de Biologie Genetică RAS, Institutul de Biologie Fizicochimică numit după V.A. Universitatea de Stat A. N. Belozersky din Moscova. Institutul de Biochimie M.V. Lomonosov. A.N. Bach RAS și Institutul de Proteine RAS din Pushchino.
Astăzi, domeniul de interes al biologilor moleculari acoperă o gamă largă de probleme științifice fundamentale. Ca și înainte, rolul principal este ocupat de studiul structurii acizilor nucleici și a biosintezei proteinelor, studiul structurii și funcțiilor diferitelor structuri intracelulare și suprafețe celulare. De asemenea, domenii importante de cercetare sunt studiul mecanismelor de recepție și transmitere a semnalului, mecanismele moleculare de transport al compușilor în interiorul celulei și, de asemenea, de la celulă la mediul extern și înapoi. Dintre principalele direcții de cercetare științifică în domeniul biologiei moleculare aplicate, una dintre cele mai prioritare este problema apariției și dezvoltării tumorilor. De asemenea, un domeniu foarte important, care este studiat de secțiunea de biologie moleculară - genetică moleculară, este studiul fundamentelor moleculare ale apariției. boli ereditare, și boli virale, precum SIDA, precum și dezvoltarea unor metode de prevenire și, eventual, de tratament a acestora la nivel de gene. Descoperirile și evoluțiile biologilor moleculari în medicina legală și-au găsit o aplicație largă. O adevărată revoluție în domeniul identificării personale a fost făcută în anii 80 de oamenii de știință din Rusia, SUA și Marea Britanie datorită dezvoltării și implementării metodei de „amprentare genomică” – identificarea ADN-ului în practica de zi cu zi. Cercetările în acest domeniu nu se opresc până în prezent, metodele moderne fac posibilă stabilirea unei persoane cu o probabilitate de eroare de o miliardime dintr-un procent. Deja, există o dezvoltare activă a proiectului unui pașaport genetic, care, așa cum era de așteptat, va reduce foarte mult nivelul criminalității.
Metodologie
Astăzi, biologia moleculară are un arsenal extins de metode pentru a rezolva cele mai avansate și mai complexe probleme cu care se confruntă oamenii de știință.
Una dintre cele mai comune metode în biologia moleculară este electroforeza pe gel, care rezolvă problema separării unui amestec de macromolecule după mărime sau sarcină. Aproape întotdeauna, după separarea macromoleculelor în gel, se folosește blotting, o metodă care vă permite să transferați macromolecule de pe gel (sorb) pe suprafața membranei pentru confortul lucrului ulterioar cu acestea, în special hibridizarea. Hibridizarea - formarea de ADN hibrid din două catene de natură diferită - o metodă care joacă un rol important în cercetarea fundamentală. Este folosit pentru a determina complementar segmente în ADN diferit (ADN de diferite specii), este folosit pentru căutarea de noi gene, interferența ARN a fost descoperită cu ajutorul său, iar principiul său a stat la baza amprentei genomice.
Un rol important în practica modernă a cercetării biologice moleculare îl joacă metoda de secvențiere - determinarea secvenței nucleotidelor din acizii nucleici și a aminoacizilor din proteine.
Biologia moleculară modernă nu poate fi imaginată fără metoda reacției în lanț a polimerazei (PCR). Datorită acestei metode, se realizează o creștere a numărului (amplificarea) de copii ale unei anumite secvențe de ADN pentru a obține dintr-o moleculă o cantitate suficientă de substanță pentru a lucra în continuare cu aceasta. Un rezultat similar este obținut prin tehnologia de clonare moleculară, în care secvența de nucleotide necesară este introdusă în ADN-ul bacteriilor (sisteme vii), după care înmulțirea bacteriilor duce la rezultatul dorit. Această abordare este mult mai complicată din punct de vedere tehnic, dar permite obținerea simultană a rezultatului exprimării secvenței de nucleotide studiate.
De asemenea, metodele de ultracentrifugare (pentru separarea macromoleculelor (cantități mari), celulelor, organitelor), microscopia electronică și fluorescentă, metodele spectrofotometrice, analiza de difracție cu raze X, autoradiografia etc. sunt utilizate pe scară largă în studiile de biologie moleculară.
Datorită progresului tehnologic și cercetării științifice în domeniul chimiei, fizicii, biologiei și informaticii, echipamentele moderne fac posibilă izolarea, studierea și modificarea genelor individuale și a proceselor în care sunt implicate.
1. Introducere.
Subiect, sarcini și metode de biologie moleculară și genetică. Semnificația geneticii „clasice” și a geneticii microorganismelor în dezvoltarea biologiei moleculare și a ingineriei genetice. Conceptul de genă în genetica „clasică” și moleculară, evoluția sa. Contribuția metodologiei ingineriei genetice la dezvoltarea geneticii moleculare. Valoarea aplicată a ingineriei genetice pentru biotehnologie.
2. Bazele moleculare ale eredității.
Conceptul de celulă, compoziția sa macromoleculară. Natura materialului genetic. Istoricul dovezilor pentru funcția genetică a ADN-ului.
2.1. Diferite tipuri de acizi nucleici. functii biologice acizi nucleici. Structura chimică, structura spațială și proprietăți fizice acizi nucleici. Caracteristicile structurale ale materialului genetic al pro- și eucariotelor. Perechi complementare de baze Watson-Crick. Cod genetic. Istoria descifrării codului genetic. Principalele proprietăți ale codului: triplet, cod fără virgule, degenerare. Caracteristici ale dicționarului de coduri, familii de codoni, codoni semantici și „fără sens”. Molecule circulare de ADN și conceptul de supraînfăşurare a ADN-ului. Topoizomerii ADN-ului și tipurile lor. Mecanisme de acțiune ale topoizomerazelor. ADN giraza bacteriană.
2.2. transcrierea ADN-ului. ARN polimeraza procariotă, subunitatea sa și structurile tridimensionale. Varietate de factori sigma. Promotor al genei procariote elemente structurale. Etapele ciclului de transcripție. Inițierea, formarea unui „complex deschis”, alungirea și terminarea transcripției. atenuarea transcripției. Reglarea expresiei operonului triptofan. „Riboswitches”. Mecanisme de terminare a transcripției. Reglarea negativă și pozitivă a transcripției. operon de lactoză. Reglarea transcripțională în dezvoltarea fagului lambda. Principiile recunoașterii ADN-ului de către proteinele reglatoare (proteina CAP și represorul fagilor lambda). Caracteristicile transcripției la eucariote. Procesarea ARN la eucariote. Capatarea, îmbinare și poliadenilare a transcrierilor. mecanisme de îmbinare. Rolul ARN nuclear mic și al factorilor proteici. Îmbinare alternativă, exemple.
2.3. Difuzare, etapele sale, funcția ribozomilor. Localizarea ribozomilor în celulă. Tipuri procariote și eucariote de ribozomi; ribozomi 70S și 80S. Morfologia ribozomilor. Împărțirea în subparticule (subunități). Legarea dependentă de codon a aminoacil-ARNt în ciclul de alungire. Interacțiunea codon-anticodon. Participarea factorului de alungire EF1 (EF-Tu) la legarea aminoacil-ARNt la ribozom. Factorul de alungire EF1B (EF-Ts), funcția sa, succesiunea de reacții cu participarea sa. Antibioticele care afectează stadiul legării dependente de codon a aminoacil-ARNt la ribozom. Antibioticele aminoglicozide (streptomicina, neomicina, kanamicina, gentamicina etc.), mecanismul lor de actiune. Tetraciclinele ca inhibitori ai legării aminoacil-ARNt la ribozom. Inițierea difuzării. Principalele etape ale procesului de inițiere. Inițierea translației la procariote: factori de inițiere, codoni inițiatori, capătul ARN 3¢ al subunității mici ribozomale și secvența Shine-Dalgarno în ARNm. Inițierea translației la eucariote: factori de inițiere, codoni inițiatori, regiune netradusă 5¢ și inițiere terminală dependentă de capac. Inițiere „internă” independentă de capac la eucariote. Transpeptidarea. Inhibitori ai transpeptidei: cloramfenicol, lincomicina, aicetină, streptogramine, anisomicină. Translocarea. Implicarea factorului de alungire EF2 (EF-G) și GTP. Inhibitori de translocație: acid fusidic, viomicina, mecanismele lor de acțiune. Terminarea traducerii. Codoni de terminare. Factori de terminare a proteinei ai procariotelor și eucariotelor; două clase de factori de terminare şi mecanisme de acţiune a acestora. Reglarea translației la procariote.
2.4. Replicarea ADN-uluiși controlul său genetic. Polimerazele implicate în replicare, caracteristicile activităților lor enzimatice. fidelitatea ADN-ului. Rolul interacțiunilor sterice între perechile de baze ADN în timpul replicării. E. coli polimerazele I, II și III. Subunitățile polimerazei III. Furcătură de replicare, fire „în frunte” și „întârziate” în timpul replicării. Fragmente din Okazaki. Complex de proteine din furca de replicare. Reglarea inițierii replicării în E. coli. Încetarea replicării în bacterii. Caracteristici ale reglării replicării plasmidelor. Replicare bidirecțională și cu inel rulant.
2.5. Recombinare, tipurile și modelele sale. Recombinare generală sau omoloagă. Rupere dublu-catenar în ADN care inițiază recombinarea. Rolul recombinării în repararea post-replicare a rupurilor duble catene. Structura de vacanță în modelul de recombinare. Enzimologia recombinării generale la E. coli. complexul RecBCD. Proteina Reca. Rolul recombinării în asigurarea sintezei ADN-ului în deteriorarea ADN-ului care întrerupe replicarea. recombinare la eucariote. Enzime de recombinare la eucariote. Recombinare specifică site-ului. Diferențele în mecanismele moleculare ale recombinării generale și specifice locului. Clasificarea recombinazelor. Tipuri de rearanjamente cromozomiale efectuate în timpul recombinării specifice locului. Rolul reglator al recombinării site-specifice în bacterii. Construcția cromozomilor eucarioți multicelulari folosind sistemul de recombinare fagică specifică locului.
2.6. Repararea ADN-ului. Clasificarea tipurilor de reparații. Repararea directă a dimerilor de timină și a guaninei metilate. Tăierea bazelor. Glicozilaze. Mecanismul de reparare a nucleotidelor nepereche (repararea nepotrivirii). Selectarea catenei de ADN care urmează să fie reparată. Reparație SOS. Proprietățile ADN polimerazelor implicate în repararea SOS la procariote și eucariote. Conceptul de " mutații adaptative” în bacterii. Repararea rupurilor duble catenare: recombinare post-replicativă omoloagă și asocierea capetelor neomologii ale moleculei de ADN. Relația dintre procesele de replicare, recombinare și reparare.
3. Procesul de mutație.
Rolul mutanților biochimici în formarea teoriei unei gene - a unei enzime. Clasificarea mutațiilor. Mutații punctuale și rearanjamente cromozomiale, mecanismul formării lor. Mutageneză spontană și indusă. Clasificarea mutagenilor. Mecanismul molecular al mutagenezei. Relația dintre mutageneză și reparare. Identificarea și selecția mutanților. Suprimare: intragenică, intergenică și fenotipică.
4. Extracromozomiale elemente genetice.
Plasmide, structura și clasificarea lor. Factorul sexual F, structura și ciclul său de viață. Rolul factorului F în mobilizarea transferului de cromozomi. Formarea donatorilor de tip Hfr și F. Mecanism de conjugare. Bacteriofage, structura și ciclul lor de viață. Bacteriofagi virulenți și temperați. Lizogenie și transducție. Transducție generală și specifică. Elemente genetice migratoare: transpozoni și secvențe IS, rolul lor în metabolismul genetic. Transpozonii ADN în genomul procariotelor și ale secvențelor eucariote a secvențelor IS-componente ale bacteriilor. ctor al bacteriilor, care determină capacitatea de a transfera materialul genetic în timpul conjugării Transpozonii bacteriilor și organismelor eucariote Mecanisme directe nereplicative și replicative ale transpozițiilor Înțelegerea transferului orizontal al transpozonilor și a rolului lor în rearanjamentele structurale (recombinarea ectopică) și în evoluția genomului
5. Studiul structurii și funcției genei.
Elemente de analiză genetică. Test de complementare cis-trans. Cartografie genetică folosind conjugare, transducție și transformare. Construirea hărților genetice. Cartografiere genetică fină. Analiza fizică a structurii genelor. analiza heteroduplex. Analiza restricțiilor. Metode de secvențiere. reacția în lanț a polimerazei. Dezvăluirea funcției unei gene.
6. Reglarea expresiei genelor. Concepte de operon și regulon. Control la nivelul inițierii transcripției. Proteine promotoare, operatore și reglatoare. Controlul pozitiv și negativ al expresiei genelor. Control la nivel de terminare a transcripției. Operoni controlați de cataboliți: modele de operoni lactoză, galactoză, arabinoză și maltoză. Operoni controlați de atenuator: un model al operonului triptofan. Reglarea multivalentă a expresiei genelor. Sisteme globale de reglementare. Răspunsul regulator la stres. control post-transcripțional. transducția semnalului. Reglarea mediată de ARN: ARN mici, ARN senzori.
7. Fundamentele ingineriei genetice. Enzime de restricție și modificări. Izolarea și clonarea genelor. Vectori pentru clonarea moleculară. Principii de construcție a ADN-ului recombinant și introducerea lor în celulele primitoare. Aspecte aplicate ale ingineriei genetice.
A). Literatura principala:
1. Watson J., Tooze J., ADN recombinant: un curs scurt. – M.: Mir, 1986.
2. Genele. – M.: Domnule. 1987.
3. Biologie moleculară: structura și biosinteza acizilor nucleici. / Ed. . - M. Şcoala superioară. 1990.
4., - Biotehnologia moleculară. M. 2002.
5. Ribozomi de spirnă și biosinteza proteinelor. – M.: facultate, 1986.
b). Literatură suplimentară:
1. Hesin al genomului. – M.: Știință. 1984.
2. Rybchin al ingineriei genetice. - Sankt Petersburg: Universitatea Tehnică de Stat din Sankt Petersburg. 1999.
3. Genele Patrushev. – M.: Nauka, 2000.
4. Microbiologie modernă. Procariote (în 2 vol.). – M.: Mir, 2005.
5. M. Singer, P. Berg. Gene și genoame. – M.: Mir, 1998.
6. ingineria Shchelkunov. - Novosibirsk: Din Sib. Univ., 2004.
7. Biologie Stepanov. Structura și funcțiile proteinelor. - M.: V. Sh., 1996.
interviu
Pirogov Sergey - un participant la pregătirea pentru Olimpiada de biologie, organizată de „Elephant and Giraffe” în 2012.
Câștigător al Universiadei Internaționale de Biologie
Câștigătorul olimpiadei „Lomonosov”
Câștigător al etapei regionale Olimpiada integrală rusească la biologie în 2012
Studiază la Universitatea de Stat din Moscova. M.V. Lomonosov la Facultatea de Biologie: Catedra de Biologie Moleculară, student în anul VI. Lucrează în Laboratorul de Genetică Biochimică a Animalelor al Institutului de Genetică Moleculară.
- Seryozha, dacă cititorii au întrebări, vă vor putea întreba?
Da, desigur, puteți pune întrebări cel puțin imediat. În acest domeniu:
Faceți clic aici pentru a pune o întrebare.

- Să începem cu școala, nu ai avut o școală super-mișto?
Am studiat la o școală foarte slabă din Moscova, un liceu atât de mediu. Adevărat, am avut un profesor minunat la Teatrul de Artă din Moscova, datorită căruia am avut o orientare în mare parte nominală de „istoria artei” a școlii.
- Dar biologie?
Profesorul nostru de biologie era o femeie foarte în vârstă, surdă și ascuțită, de care toată lumea se temea. Dar dragostea pentru subiectul ei nu a adăugat. Sunt pasionat de biologie încă din copilărie, de la vârsta de cinci ani. Am citit totul singur, fiind purtată în principal de anatomie și zoologie. Asa de rechizite exista paralel cu propriile mele interese. Jocurile Olimpice au schimbat totul.
- Spune-mi mai mult despre aceasta.
În clasa a VII-a am participat pentru prima dată la etapa municipală (desigur, la aproape toate materiile deodată, fiind singurul elev pe care profesorii aveau de ce să-l trimită). Și a câștigat la biologie. Apoi școala a tratat acest lucru ca pe un fapt amuzant, dar nu foarte interesant.

- Te-a ajutat la școală?
Îmi amintesc că, în ciuda studiilor mele strălucitoare, am primit adesea B de la un profesor de biologie cu strângere de niște de genul „în desenul unei secțiuni de ceapă, rădăcinile ar trebui să fie vopsite maro, nu gri”. Totul a fost destul de deprimant. În clasa a VIII-a am fost din nou la olimpiada, dar din anumite motive nu am fost trimis la biologie. Dar a devenit câștigător și premiat la alte materii.
- Ce s-a întâmplat în clasa a IX-a?
În clasa a IX-a nu am mers la etapa raională. Acolo am înscris în mod neașteptat un scor slab, la limită, care, totuși, s-a dovedit a trece la etapa regională. A avut o forță de motivare puternică - realizarea cât de multe nu știu și câți oameni care știu toate acestea (câți astfel de oameni la scară națională chiar îmi era teamă să-mi imaginez).

- Spune-ne cum te-ai pregătit.
Studiul individual intensiv, incursiunile în librării și mii de sarcini de anul trecut au avut un efect vindecător. Am obținut unul dintre cele mai mari scoruri la teorie (care a fost și pentru mine complet neașteptat), am trecut la etapa practică... și am eșuat. Pe vremea aceea, nici măcar nu știam de existența etapei practice.
- Te-au influențat Olimpiada?
Viața mea s-a schimbat radical. Am aflat despre multe alte olimpiade, în special m-am îndrăgostit de SBO. Ulterior, a arătat multe rezultate bune, unii au câștigat, datorită lui „Lomonosovskaya” au primit dreptul de a intra fără examene. În același timp, am câștigat olimpiade în istoria artei, la care încă respir inegal. Adevărat, nu era prieten cu tururi practice. În clasa a XI-a am ajuns totuși la etapa finală, dar Fortune nu a fost favorabil, iar de data aceasta nu am avut timp să completez matricea de răspuns a etapei teoretice. Dar acest lucru a făcut posibil să nu vă faceți prea multe griji cu privire la practic.
- Ai întâlnit multe olimpiade?
Da, mai cred că am avut mare noroc cu cercul semenilor mei, care mi-au lărgit foarte mult orizonturile. Cealaltă parte a olimpiadelor, pe lângă motivația de a studia subiectul mai armonios, a fost cunoașterea olimpiadelor. Deja la vremea respectivă, am observat că comunicarea orizontală este uneori mai utilă decât cea verticală - cu profesorii în cantonament.

- Cum ai intrat la universitate? Ai ales o facultate?
După clasa a XI-a, am intrat la Facultatea de Biologie a Universității de Stat din Moscova. Doar majoritatea camarazilor mei de atunci au făcut o alegere în favoarea FBB, dar aici rolul principal a fost jucat de faptul că nu am devenit câștigătorul All-Russian. Așa că ar trebui să dau un examen intern la matematică, iar în el, mai ales la școală - m-am îndrăgostit mult mai mult de cel superior - nu eram puternic. Și a fost o pregătire foarte slabă la școală (nu eram pregătiți pentru aproape toată partea C). La capitolul interese, chiar și atunci am bănuit că, până la urmă, poți ajunge la orice rezultat, indiferent de locul de admitere. Ulterior, s-a dovedit că există mulți absolvenți FBB care au trecut la biologie predominant umedă și invers - mulți bioinformaticieni buni au început ca amatori. Deși în acel moment mi se părea că contingentul de la facultatea de biologică va fi diferit de cel FBBshny. În asta cu siguranță m-am înșelat.
Știați?
Interesant
Știați?
Interesant
În tabăra Elefant și Girafă au loc schimbări în biochimie și biologie moleculară, unde școlari, împreună cu profesori cu experiență de la Universitatea de Stat din Moscova, organizează experimente și se pregătesc și pentru olimpiade.© Intervievat de Reshetov Denis. Fotografiile au fost oferite cu amabilitate de Sergey Pirogov.
Progresele în studiul acizilor nucleici și al biosintezei proteinelor au condus la crearea unui număr de metode de mare importanță practică în medicină, agricultură și o serie de alte industrii.
După ce au fost studiate codul genetic și principiile de bază ale stocării și implementării informațiilor ereditare, dezvoltarea biologiei moleculare a ajuns într-un impas, deoarece nu existau metode care să facă posibilă manipularea genelor, izolarea și schimbarea acestora. Apariția acestor metode a avut loc în anii 1970-1980. Aceasta a dat un impuls puternic dezvoltării acestui domeniu al științei, care înflorește și astăzi. În primul rând, aceste metode vizează obținerea de gene individuale și introducerea lor în celulele altor organisme (clonarea moleculară și transgeneza, PCR), precum și metodele de determinare a secvenței nucleotidelor din gene (secvențierea ADN și ARN). Aceste metode vor fi discutate mai detaliat mai jos. Vom începe cu cea mai simplă metodă de bază, electroforeza, apoi vom trece la metode mai complexe.
ELECTROFOREZA ADN-ului
Este metoda de bază de lucru cu ADN, care este folosită împreună cu aproape toate celelalte metode pentru a izola moleculele dorite și a analiza rezultatele. Electroforeza pe gel este utilizată pentru a separa fragmentele de ADN după lungime. ADN-ul este un acid, moleculele sale conțin reziduuri de acid fosforic, care desprind un proton și capătă o sarcină negativă (Fig. 1).
Prin urmare, într-un câmp electric, moleculele de ADN se deplasează spre anod - un electrod încărcat pozitiv. Acest lucru se întâmplă într-o soluție de electrolit care conține ioni purtători de sarcină, datorită cărora această soluție conduce curentul. Pentru a separa fragmentele, se folosește un gel dens din polimeri (agaroză sau poliacrilamidă). Moleculele de ADN se „încurcă” în ea cu atât mai mult, cu atât sunt mai lungi și, prin urmare, cele mai lungi molecule se mișcă cel mai încet, iar cele mai scurte - cel mai rapid (Fig. 2). Înainte sau după electroforeză, gelul este tratat cu coloranți care se leagă de ADN și fluoresc în lumină ultravioletă și se obține un model de benzi în gel (vezi Fig. 3). Pentru a determina lungimile fragmentelor de ADN dintr-o probă, acestea sunt comparate cu un marker, adică un set de fragmente de lungimi standard depuse în paralel pe același gel (Fig. 4).



Cele mai importante instrumente pentru lucrul cu ADN-ul sunt enzimele care efectuează transformări ADN-ului în celulele vii: ADN polimeraze, ADN ligaze și endonucleaze de restricție sau enzime de restricție. ADN polimeraza Se realizează sinteza șablonului ADN, care permite propagarea ADN-ului într-o eprubetă. ADN ligaze coase moleculele de ADN împreună sau vindecă golurile din ele. Endonucleaze de restricție, sau restrictaze, tăiați moleculele de ADN în funcție de secvențe strict definite, ceea ce vă permite să decupați fragmente individuale din masa totală de ADN. Aceste fragmente pot conține în unele cazuri gene individuale.
restrictaze
Secvențele recunoscute de enzimele de restricție sunt simetrice, iar rupturile pot apărea la mijlocul unei astfel de secvențe sau cu o deplasare (în același loc în ambele catene de ADN). Schema de actiune tipuri diferite restrictaza este prezentată în fig. 1. În primul caz se obțin așa-numitele capete „tocite”, iar în al doilea - capete „lipioase”. În cazul capetelor „lipicioase” ale fundului, lanțul este mai scurt decât celălalt, se formează o secțiune monocatenară cu o secvență simetrică care este aceeași la ambele capete formate.
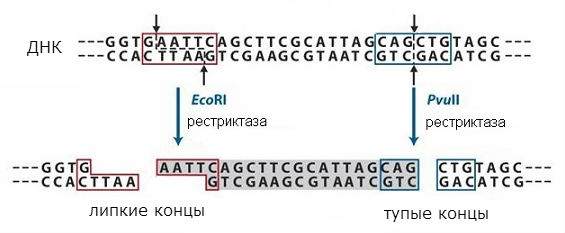
Secvențele finale vor fi aceleași atunci când orice ADN este scindat cu o enzimă de restricție dată și pot fi reunite deoarece au secvențe complementare. Pot fi ligați cu ADN ligază pentru a forma o singură moleculă. Astfel, este posibil să combinați fragmente din două ADN diferite și să obțineți așa-numitul ADN recombinat. Această abordare este utilizată în metoda clonării moleculare, care face posibilă obținerea de gene individuale și introducerea lor în celule care pot forma proteina codificată în genă.
clonarea moleculară
Clonarea moleculară folosește două molecule de ADN - o inserție care conține gena de interes și vector- ADN care acționează ca purtător. Insertul este „cusut” in vector cu ajutorul enzimelor, obtinandu-se o noua molecula de ADN recombinant, apoi aceasta molecula este introdusa in celulele gazda, iar aceste celule formeaza colonii pe un mediu nutritiv. O colonie este descendentul unei celule, adică o clonă, toate celulele coloniei sunt identice genetic și conțin același ADN recombinant. De aici și termenul de „clonare moleculară”, adică obținerea unei clone de celule care conține un fragment de ADN de interes pentru noi. După ce sunt obținute coloniile care conțin insertul care ne interesează, este posibil să se caracterizeze acest insert prin diferite metode, de exemplu, pentru a determina secvența exactă a acestuia. Celulele pot produce, de asemenea, proteina codificată de insert dacă aceasta conține o genă funcțională.
Când o moleculă recombinantă este introdusă în celule, are loc transformarea genetică a acestor celule. Transformare- procesul de absorbție de către o celulă a unui organism a unei molecule de ADN liber din mediu și integrarea acesteia în genom, ceea ce duce la apariția într-o astfel de celulă a unor noi trăsături ereditare pentru aceasta, caracteristice organismului-donator de ADN. De exemplu, dacă molecula introdusă conține o genă de rezistență la antibioticul ampicilină, atunci bacteriile transformate vor crește în prezența acesteia. Înainte de transformare, ampicilina le-a provocat moartea, adică un nou semn apare în celulele transformate.
VECTORI
Un vector trebuie să aibă un număr de proprietăți:
În primul rând, este o moleculă de ADN relativ mică, care poate fi manipulată cu ușurință.
În al doilea rând, pentru ca ADN-ul să fie conservat și reprodus într-o celulă, acesta trebuie să conțină o anumită secvență care să îi asigure replicarea (originea replicării, sau originea replicării).
În al treilea rând, trebuie să conțină gena marker, care asigură selecția doar a acelor celule în care a intrat vectorul. De obicei, acestea sunt gene de rezistență la antibiotice - apoi, în prezența unui antibiotic, toate celulele care nu conțin vectorul mor.
Clonarea genelor se realizează cel mai adesea în celulele bacteriene, deoarece acestea sunt ușor de cultivat și se înmulțesc rapid. Într-o celulă bacteriană, există de obicei o moleculă circulară mare de ADN, lungă de câteva milioane de perechi de baze, care conține toate genele necesare bacteriilor - cromozomul bacterian. Pe lângă aceasta, în unele bacterii există mici (câteva mii de perechi de baze) ADN circular, numit plasmide(Fig. 2). Ele, ca și ADN-ul principal, conțin o secvență de nucleotide care oferă capacitatea ADN-ului de a se replica (ori). Plasmidele se replic independent de ADN-ul principal (cromozomial), prin urmare sunt prezente în celula din în număr mare copii. Multe dintre aceste plasmide poartă gene de rezistență la antibiotice, ceea ce face posibilă distingerea celulelor care poartă plasmida de celulele normale. Mai frecvent, sunt utilizate plasmide care poartă două gene care conferă rezistență la două antibiotice, cum ar fi tetraciclina și amicilina. Exista metode simple izolarea unui astfel de ADN plasmidic liber de ADN-ul cromozomului principal al bacteriei.

SEMNIFICAȚIA TRANSGENEZEI
Se numește transferul de gene de la un organism la altul transgenezași astfel de organisme modificate - transgenic. Metoda de transfer de gene în celulele microbiene este utilizată pentru a obține preparate de proteine recombinante pentru medicină, în special, proteine umane care nu provoacă respingere imună - interferoni, insulină și alți hormoni proteici, factori de creștere celulară, precum și proteine pentru producerea de vaccinuri. În cazuri mai complexe, când modificarea proteinelor este efectuată corect numai în celulele eucariote, se folosesc culturi de celule transgenice sau animale transgenice, în special, animale (în primul rând capre), care secretă proteinele necesare în lapte, sau proteinele sunt izolate din sângele lor. Așa se obțin anticorpii, factorii de coagulare a sângelui și alte proteine. obţinut prin transgeneză plante cultivate care sunt rezistente la erbicide și dăunători și au alte proprietăți benefice. Folosind microorganisme transgenice pentru a purifica apele uzate și a lupta împotriva poluării, există chiar și microbi transgenici care pot descompune petrolul. În plus, tehnologiile transgenice sunt indispensabile în cercetare științifică- dezvoltarea biologiei de astăzi este de neconceput fără utilizarea de rutină a metodelor de modificare și transfer de gene.
tehnologie de clonare moleculară
inserții
Pentru a obține o genă individuală din orice organism, tot ADN-ul cromozomial este izolat din acesta și scindat cu una sau două enzime de restricție. Enzimele sunt selectate astfel încât să nu taie gena care ne interesează, ci să facă rupturi de-a lungul marginilor ei, iar în ADN-ul plasmidic să facă o ruptură într-una dintre genele de rezistență, de exemplu, la ampicilină.
Procesul de clonare moleculară include următorii pași:
Tăiere și cusătură - construcția unei singure molecule recombinante dintr-o inserție și un vector.
Transformarea este introducerea unei molecule recombinante în celule.
Selecție - selecție de celule care au primit un vector cu o inserție.
taierea si cusatura
ADN-ul plasmidic este tratat cu aceleași enzime de restricție și se transformă într-o moleculă liniară dacă este selectată o astfel de enzimă de restricție care introduce 1 ruptură în plasmidă. Ca urmare, aceleași capete lipicioase apar la capetele tuturor fragmentelor de ADN rezultate. Pe măsură ce temperatura scade, aceste capete se unesc aleatoriu și sunt legate cu ADN ligază (vezi Fig. 3).

Se obține un amestec de ADN-uri circulare de compoziție diferită: unele dintre ele vor conține o anumită secvență de ADN cromozomială conectată la ADN-ul bacterian, altele vor conține fragmente de ADN cromozomial unite între ele, iar altele vor conține o plasmidă circulară redusă sau dimerul acesteia (Fig. 4).

transformare
În continuare, se efectuează acest amestec transformare genetică bacterii care nu contin plasmide. Transformare- procesul de absorbție de către o celulă a unui organism a unei molecule de ADN liber din mediu și integrarea acesteia în genom, ceea ce duce la apariția într-o astfel de celulă a unor noi trăsături ereditare pentru aceasta, caracteristice organismului-donator de ADN. Doar o plasmidă poate intra și se poate multiplica în fiecare celulă. Astfel de celule sunt plasate pe un mediu nutritiv solid care conține antibioticul tetraciclină. Celulele care nu au primit plasmida nu vor crește pe acest mediu, iar celulele care poartă plasmida formează colonii, fiecare dintre acestea conținând descendenții unei singure celule, adică. toate celulele dintr-o colonie poartă aceeași plasmidă (vezi Fig. 5).

Selecţie
În continuare, sarcina este de a izola numai celulele în care a intrat vectorul cu inserția și de a le distinge de celulele care poartă doar vectorul fără inserție sau care nu poartă vectorul deloc. Acest proces de selectare a celulelor potrivite este numit selecţie. Pentru aceasta, aplicați markeri selectivi- de obicei gene de rezistență la antibiotice în vector, și medii selective care conțin antibiotice sau alte substanțe selective.
În exemplul pe care îl luăm în considerare, celulele din coloniile crescute în prezența ampicilinei sunt subcultivate pe două medii: primul conține ampicilină, iar al doilea conține tetraciclină. Coloniile care conțin doar plasmida vor crește pe ambele medii, în timp ce coloniile care conțin ADN cromozomial inserat în plasmide nu vor crește pe mediul cu tetraciclină (Fig. 5). Dintre acestea, cele care conțin gena care ne interesează sunt selectate prin metode speciale, crescute în cantități suficiente, iar ADN-ul plasmidic este izolat. Din acesta, folosind aceleași restrictaze care au fost folosite pentru a obține ADN recombinant, gena individuală de interes este tăiată. ADN-ul acestei gene poate fi folosit pentru a determina secvența de nucleotide, poate fi introdus în orice organism pentru a obține noi proprietăți sau pentru a sintetiza proteina dorită. Această metodă de izolare a genelor se numește clonarea moleculară.
PROTEINE FLUORESCENTE
Este foarte convenabil să folosiți proteine fluorescente ca gene marker în studiile organismelor eucariote. Gena pentru prima proteină fluorescentă, proteină verde fluorescentă (GFP) a fost izolat din meduza Aqeuorea victoria și introdus în diferite organisme model (vezi Fig. 6) Premiul Nobel pentru descoperirea și utilizarea acestei proteine.

Apoi au fost izolate genele pentru alte proteine fluorescente - roșu, albastru, galben. Aceste gene au fost modificate artificial pentru a produce proteine cu proprietățile dorite. Diversitatea proteinelor fluorescente este prezentată în fig. 7, care prezintă o placă Petri cu bacterii care conțin gene pentru diferite proteine fluorescente.

aplicarea proteinelor fluorescente
Gena proteinei fluorescente poate fi fuzionată cu gena oricărei alte proteine, apoi în timpul translației se va forma o singură proteină - o proteină de fuziune translațională sau fuziune(proteina de fuziune), care are fluorescență. Astfel, este posibil să se studieze, de exemplu, localizarea (locația) oricăror proteine de interes în celulă, mișcarea acestora. Exprimând proteine fluorescente numai în anumite tipuri de celule, este posibil să se eticheteze celulele de aceste tipuri în organism pluricelular(vezi Fig. 8 - creierul de șoarece, în care neuronii individuali au culori diferite datorită unei anumite combinații de gene proteice fluorescente). Proteinele fluorescente sunt un instrument indispensabil în biologia moleculară modernă.

PCR
O altă metodă de obținere a genelor se numește reacția în lanț a polimerazei (PCR). Se bazează pe capacitatea ADN polimerazelor de a completa a doua catenă de ADN de-a lungul catenei complementare, așa cum se întâmplă în celule în timpul replicării ADN-ului.
Originile replicării în această metodă sunt date de două bucăți mici de ADN numite seminte, sau grunduri. Acești primeri sunt complementari capetelor genei de interes de pe două catene de ADN. În primul rând, ADN-ul cromozomial din care urmează să fie izolată gena este amestecat cu semințe și încălzit la 99 ° C. Acest lucru duce la ruperea legăturilor de hidrogen și la divergența catenelor de ADN. După aceea, temperatura este coborâtă la 50-70 aproximativ C (în funcție de lungimea și succesiunea semințelor). În aceste condiții, primerii sunt atașați de regiuni complementare ale ADN-ului cromozomial, formând o dublă helix regulată (vezi Fig. 9). După aceea, se adaugă un amestec din toate cele patru nucleotide necesare pentru sinteza ADN-ului și ADN polimeraza. Enzima alungește primerii prin construirea de ADN dublu catenar din punctul de atașare a primerilor, adică. de la capetele unei gene până la capătul unei molecule de cromozom monocatenar. 
Dacă amestecul este acum încălzit din nou, lanțurile cromozomiale și cele nou sintetizate se vor dispersa. După răcire, li se vor alătura din nou semințele, care sunt luate în exces mare (vezi Fig. 10).

Pe lanțurile nou sintetizate, acestea se vor uni nu la capătul de la care a început prima sinteză, ci la cel opus, deoarece lanțurile de ADN sunt antiparalele. Prin urmare, în al doilea ciclu de sinteză, numai secvența corespunzătoare genei va fi completată pe astfel de lanțuri (vezi Fig. 11).
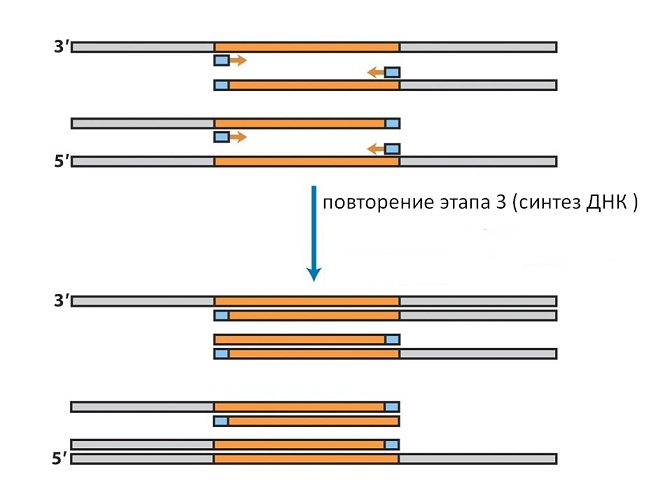
Această metodă utilizează ADN polimeraza din bacterii termofile care pot rezista la fierbere și funcționează la temperaturi de 70-80 ° C, nu trebuie adăugată de fiecare dată, dar este suficient să o adăugați la începutul experimentului. Repetând procedurile de încălzire și răcire în aceeași secvență, putem dubla numărul de secvențe din fiecare ciclu, delimitat la ambele capete de semințele introduse (vezi Fig. 12).

După aproximativ 25 de astfel de cicluri, numărul de copii ale genei va crește de peste un milion de ori. Astfel de cantități pot fi ușor separate de ADN-ul cromozomial introdus în eprubetă și utilizate în diverse scopuri.
Secvențierea ADN-ului
O altă realizare importantă este dezvoltarea metodelor de determinare a secvenței nucleotidelor din ADN - Secvențierea ADN-ului(din engleză sequence - sequence). Pentru a face acest lucru, este necesar să obțineți gene pure din alt ADN folosind una dintre metodele descrise. Apoi lanțurile de ADN sunt separate prin încălzire și li se adaugă un primer marcat cu fosfor radioactiv sau o etichetă fluorescentă. Vă rugăm să rețineți că se ia o sămânță, complementară unui lanț. Apoi se adaugă ADN polimeraza și un amestec de 4 nucleotide. Un astfel de amestec este împărțit în 4 părți și la fiecare se adaugă una dintre nucleotide, modificată astfel încât să nu conțină o grupare hidroxil pe al treilea atom de deoxiriboză. Dacă o astfel de nucleotidă este inclusă în lanțul de ADN sintetizat, atunci alungirea sa nu va putea continua, deoarece polimeraza nu va avea de unde să atașeze următoarea nucleotidă. Prin urmare, sinteza ADN-ului după includerea unei astfel de nucleotide este întreruptă. Aceste nucleotide, numite dideoxinucleotide, sunt adăugate mult mai puțin decât de obicei, astfel încât terminarea lanțului are loc doar ocazional și în fiecare lanț în locuri diferite. Rezultatul este un amestec de lanțuri de lungimi diferite, fiecare având aceeași nucleotidă la capăt. Astfel, lungimea lanțului corespunde numărului de nucleotide din secvența studiată, de exemplu, dacă am avut o adenil dideoxinucleotidă, iar lanțurile rezultate aveau lungimea de 2, 7 și 12 nucleotide, atunci adenina se afla în pozițiile a doua, a șaptea și a douăsprezecea în genă. Amestecul rezultat de lanțuri poate fi ușor separat după dimensiune folosind electroforeză, iar lanțurile sintetizate pot fi identificate prin radioactivitate pe film cu raze X (vezi Fig. 10).

Se pare că imaginea din partea de jos a imaginii, numită radioautograf. Deplasându-ne de jos în sus și citind litera de deasupra coloanelor fiecărei zone, vom obține secvența de nucleotide prezentată în figura din dreapta autografului. S-a dovedit că sinteza este oprită nu numai de dideoxinucleotide, ci și de nucleotide, în care un fel de grup chimic cum ar fi un colorant fluorescent. Dacă fiecare nucleotidă este marcată cu propriul colorant, atunci zonele obținute prin separarea lanțurilor sintetizate vor străluci cu o lumină diferită. Acest lucru face posibilă efectuarea reacției într-o eprubetă simultan pentru toate nucleotidele și, prin separarea lanțurilor rezultate după lungime, identificarea nucleotidelor după culoare (vezi Fig. 11).

Astfel de metode au făcut posibilă determinarea secvențelor nu numai ale genelor individuale, ci și citirea genomilor întregi. Au fost dezvoltate acum metode și mai rapide pentru determinarea secvențelor de nucleotide din gene. Dacă primul genom uman a fost descifrat de un mare consorțiu internațional folosind prima metodă dată în 12 ani, a doua, folosind a doua, în trei ani, acum acest lucru se poate face într-o lună. Acest lucru vă permite să preziceți predispoziția unei persoane la multe boli și să luați măsuri în avans pentru a le evita.




