La descifrarea genomului insectei s-a constatat că. Genom într-un genom: un parazit vechi. Dificultăți în studierea genomului plantelor
La 50 de ani de la descoperirea structurii ADN-ului
A.V. Zelenin
GENOMUL PLANTELOR
A. V. Zelenin
Zelenin Alexandru Vladimirovici- d.b.n.,
şef al laboratorului Institutului de Biologie Moleculară. V.A. Engelhardt RAS.
Realizările impresionante ale programului „Genom uman”, precum și succesul lucrărilor de descifrare a așa-numitelor genomuri extra-mici (virusuri), mici (bacterii, drojdie) și medii (viermi rotunzi, Drosophila), au făcut posibilă trecerea la un studiu pe scară largă a genomului plantelor mari și foarte mari. Necesitatea urgentă a unui studiu detaliat al genomilor celor mai importante plante din punct de vedere economic a fost subliniată la o întâlnire despre genomica plantelor, organizată în 1997 în Statele Unite [ , ]. De-a lungul anilor care au trecut de atunci, s-au obținut succese neîndoielnice în acest domeniu. În 2000, a apărut o publicație privind secvențierea completă (determinarea secvenței nucleotidice liniare a întregului ADN nuclear) a genomului muștarului mic - Arabidopsis, în 2001 - privind secvențierea preliminară (proiect) a genomului orezului. Au fost raportate în mod repetat lucrări de secvențiere a genomului plantelor mari și super-mari (porumb, secară, grâu), cu toate acestea, aceste rapoarte nu conțineau informații specifice și erau mai degrabă de natura declarațiilor de intenție.
Se presupune că decodificarea genomului plantelor va deschide perspective largi pentru știință și practică. În primul rând, identificarea de noi gene și lanțul de reglare genetică a acestora va crește semnificativ productivitatea plantelor prin utilizarea abordărilor biotehnologice. Odată cu descoperirea, izolarea, reproducerea (clonarea) și secvențierea genelor responsabile pentru funcții atât de importante ale organismului vegetal precum reproducerea și productivitatea, procesele de variabilitate, rezistența la factorii de mediu adversi, precum și împerecherea omoloagă a cromozomilor, apariția se asociază noi oportunităţi de îmbunătăţire a procesului de ameliorare . În cele din urmă, genele izolate și donate pot fi folosite pentru a obține plante transgenice cu proprietăți fundamental noi și pentru a analiza mecanismele de reglare a activității genelor.
Importanța studierii genomului plantelor este subliniată și de faptul că până în prezent numărul de gene de plante localizate, clonate și secvențiate este mic și variază, după diverse estimări, între 800 și 1200. Acesta este de 10-15 ori mai mic decât, pt. exemplu, la oameni.
Statele Unite rămân liderul incontestabil în studiul pe scară largă a genomului plantelor, deși studii intensive ale genomului orezului sunt efectuate în Japonia și în anul trecutși în China. La descifrarea genomului Arabidopsis, pe lângă laboratoarele din SUA, au participat activ grupurile de cercetare europene. Conducerea aparentă a Statelor Unite provoacă îngrijorare serioasă a oamenilor de știință europeni, pe care aceștia au exprimat-o în mod clar la o întâlnire sub titlul semnificativ „Perspective pentru genomica în era post-genomică”, desfășurată la sfârșitul anului 2000 în Franța. Progresul științei americane în studierea genomului plantelor agricole și crearea formelor de plante transgenice, potrivit oamenilor de știință europeni, amenință că într-un viitor nu prea îndepărtat (două-cinci decenii), când creșterea populației va pune omenirea în fața unui criza alimentară, economia și știința europeană vor deveni dependente de tehnologia americană. În acest sens, a fost anunțată crearea unui program științific franco-german pentru studiul genomului plantelor („Plantgene”) și s-au făcut investiții semnificative în acesta.
Evident, problemele genomicei plantelor ar trebui să atragă atenția oamenilor de știință și organizatorilor ruși ai științei, precum și a organelor de conducere, deoarece nu este vorba doar de prestigiul științific, ci și de securitatea națională a țării. Într-un deceniu sau două, alimentele vor deveni cea mai importantă resursă strategică.
DIFICULTĂȚI ÎN STUDIAREA GENOMULUI PLANTELOR
Studiul genomului plantelor este o sarcină mult mai dificilă decât studiul genomului uman și al altor animale. Acest lucru se datorează următoarelor circumstanțe:
dimensiuni uriașe ale genomului, atingând zeci și chiar sute de miliarde de perechi de baze (pb) pentru speciile individuale de plante: genomul principalelor plante importante din punct de vedere economic (cu excepția orezului, inului și bumbacului) fie sunt apropiate ca mărime de genomul uman, fie depășește-l de multe ori (tabel);STUDII CROMOZOMIALE ALE GENOMURILORFluctuații puternice ale numărului de cromozomi la diferite plante - de la două la unele specii la câteva sute la altele și nu este posibil să se identifice o corelație strictă între dimensiunea genomului și numărul de cromozomi;
Se formează o abundență de poliploide (conținând mai mult de doi genomi per celulă) cu genomi similari, dar nu identici (alpoliploidie);
Îmbogățirea extremă a genomului plantelor (până la 99%) de ADN „nesemnificativ” (necodificare, adică neconținând gene), ceea ce face foarte dificilă unirea (aranjarea în ordinea corectă) a fragmentelor secvențiate într-un mod comun. regiune ADN de dimensiuni mari (contig);
Cartografierea morfologică, genetică și fizică incompletă a cromozomilor (comparativ cu genomul Drosophila, uman și șoarece);
Imposibilitatea practică a izolării cromozomilor individuali în formă pură folosind metode utilizate de obicei în acest scop pentru cromozomi umani și animale (sortarea în flux și utilizarea hibrizilor celulari);
Dificultatea cartografierii cromozomiale (determinarea locației pe cromozom) a genelor individuale folosind hibridizare in situ datorită atât conținutului ridicat de ADN „nesemnificativ” din genomul plantelor, cât și particularităților organizarea structurală cromozomi vegetali;
Depărtarea evolutivă a plantelor față de animale, ceea ce complică serios utilizarea informațiilor obținute prin secvențierea genomului oamenilor și a altor animale pentru studiul genomului plantelor;
Procesul lung de reproducere al majorității plantelor, care încetinește semnificativ analiza genetică a acestora.
Studiile cromozomiale (citogenetice) ale genomului în general și ale plantelor în special au o istorie lungă. Termenul „genom” a fost propus pentru a desemna un set haploid (unic) de cromozomi cu genele conținute în ei în primul sfert al secolului al XX-lea, adică cu mult înainte de stabilirea rolului ADN-ului ca purtător de informații genetice. .
Descrierea genomului unui organism multicelular nou, nestudiat genetic anterior, începe de obicei cu studiul și descrierea setului complet al cromozomilor săi (cariotip). Acest lucru, desigur, se aplică și plantelor, dintre care un număr mare nici măcar nu au început să fie studiate.
Deja în zorii studiilor cromozomiale, genomurile speciilor de plante înrudite au fost comparate pe baza analizei conjugării meiotice (combinație de cromozomi omologi) în hibrizi interspecifici. În ultimii 100 de ani, posibilitățile de analiză a cromozomilor s-au extins dramatic. Acum, tehnologii mai avansate sunt folosite pentru a caracteriza genomul plantelor: diverse variante ale așa-numitei colorări diferențiale, care permite caracteristici morfologice identificarea cromozomilor individuali; hibridizare in situ făcând posibilă localizarea unor gene specifice pe cromozomi; studii biochimice ale proteinelor celulare (electroforeza si imunochimie) si, in final, un set de metode bazate pe analiza ADN-ului cromozomial pana la secventierea acestuia.

Orez. 1. Cariotipurile cerealelor a - secară (14 cromozomi), b - grâu dur (28 cromozomi), c - grâu moale (42 cromozomi), d - orz (14 cromozomi)De mulți ani, au fost studiate cariotipurile cerealelor, în primul rând grâul și secară. Interesant este că la diferite specii ale acestor plante, numărul de cromozomi este diferit, dar întotdeauna un multiplu de șapte. Tipurile individuale de cereale pot fi recunoscute în mod fiabil după cariotipul lor. De exemplu, genomul de secară este format din șapte perechi de cromozomi mari cu blocuri heterocromatice intens colorate la capete, adesea numite segmente sau benzi (Fig. 1a). Genomul grâului are deja 14 și 21 de perechi de cromozomi (Fig. 1, b, c), iar distribuția blocurilor heterocromatice în ele nu este aceeași ca și în cromozomii de secară. Genomii individuali de grâu, desemnați A, B și D, diferă, de asemenea, unul de celălalt. O creștere a numărului de cromozomi de la 14 la 21 duce la o schimbare bruscă a proprietăților grâului, care se reflectă în numele lor: durum, sau paste, grâu și moale, sau pâine, grâu . Gena D, care conține gene pentru proteinele glutenului, care conferă aluatului așa-numita germinație, este responsabilă pentru dobândirea de înalte proprietăți de coacere de către grâul moale. Acest genom este căruia i se acordă o atenție deosebită în îmbunătățirea selecției grâului de pâine. O altă cereală cu 14 cromozomi, orzul (Fig. 1, d), nu este de obicei folosită pentru a face pâine, dar este principala materie primă pentru fabricarea unor produse atât de comune precum berea și whisky-ul.
Cromozomii unor plante sălbatice utilizate pentru îmbunătățirea calității celor mai importante specii agricole, precum rudele sălbatice ale grâului - Aegilops, sunt în curs de studiu intens. Noi forme de plante sunt create prin încrucișare (Fig. 2) și selecție. În ultimii ani, o îmbunătățire semnificativă a metodelor de cercetare a făcut posibilă începerea studiului genomului plantelor, ale căror cariotipuri (în principal dimensiunea mică a cromozomilor) le făceau anterior inaccesibile pentru analiza cromozomilor. Așadar, abia recent au fost identificați pentru prima dată toți cromozomii de bumbac, mușețel și in.

Orez. 2. Cariotipuri de grâu și un hibrid de grâu cu Aegilops
a - grâu moale hexaploid ( Triticum astivum), constând din genomi A, B și O; b - grâu tetraploid ( Triticum timopheevi), constând din genomi A și G. conține gene de rezistență la majoritatea bolilor grâului; c - hibrizi Triticum astivum X Triticum timopheevi rezistent la mucegai și rugină, înlocuirea unei părți a cromozomilor este clar vizibilăSTRUCTURA PRIMARĂ A ADN-ului
Odată cu dezvoltarea geneticii moleculare, însuși conceptul de genom s-a extins. Acum acest termen este interpretat atât în sensul cromozomial clasic, cât și în sensul molecular modern: întregul material genetic al unui virus, al unei celule și al unui organism individual. Desigur, în urma studiului completului structura primara genomi (aceasta este adesea numită secvența liniară completă a bazelor acizi nucleici) a unui număr de microorganisme și oameni, s-a pus problema secvențierii genomului plantelor.
Dintre numeroasele organisme vegetale, două au fost selectate pentru studiu - Arabidopsis, reprezentând clasa dicotiledoneelor (dimensiunea genomului 125 milioane bp) și orez din clasa monocotiledonelor (420-470 milioane bp). Acești genomi sunt mici în comparație cu alți genomi de plante și conțin relativ puține segmente ADN repetitive. Astfel de caracteristici au dat speranța că genomii selectați vor fi disponibili pentru determinarea relativ rapidă a structurii lor primare.

Orez. 3. Arabidopsis - muștar mic - o plantă mică din familia cruciferelor ( Brassicaceae). Pe un spațiu egal ca suprafață cu o pagină a revistei noastre, puteți crește până la o mie de organisme Arabidopsis individuale.Motivul alegerii Arabidopsis nu a fost doar dimensiunea redusă a genomului său, ci și dimensiunea redusă a organismului, ceea ce face ușoară creșterea lui în laborator (Fig. 3). Am luat în considerare ciclul său scurt de reproducere, datorită căruia este posibil să se efectueze rapid experimente privind încrucișarea și selecția, genetica studiată în detaliu, ușurința manipulării în condițiile de creștere în schimbare (modificări ale compoziției de sare a solului, adăugarea de diferite nutrienți etc.) și testarea efectului diverșilor factori mutageni și agenți patogeni (viruși, bacterii, ciuperci) asupra plantelor. Arabidopsis nu are valoare economică, prin urmare, genomul său, împreună cu genomul șoarecelui, a fost numit referință sau, mai puțin precis, model.*
* Apariția termenului „genom model” în literatura rusă este rezultatul unei traduceri inexacte a expresiei engleze model genom. Cuvântul „model” înseamnă nu numai adjectivul „model”, ci și substantivul „probă”, „standard”, „model”. Ar fi mai corect să vorbim despre un genom eșantion sau despre un genom de referință.Lucrările intensive privind secvențierea genomului Arabidopsis au fost începute în 1996 de către un consorțiu internațional care includea instituții științifice și grupuri de cercetare din SUA, Japonia, Belgia, Italia, Marea Britanie și Germania. În decembrie 2000, au devenit disponibile informații extinse care rezumă determinarea structurii primare a genomului Arabidopsis. Pentru secvențiere a fost folosită tehnologia clasică sau ierarhică: mai întâi, au fost studiate secțiuni individuale mici ale genomului, din care au fost compuse secțiuni mai mari (contigs), iar în etapa finală, structura cromozomilor individuali. ADN-ul nuclear al genomului Arabidopsis este distribuit pe cinci cromozomi. În 1999, au fost publicate rezultatele secvențierii a doi cromozomi, iar apariția în presă a informațiilor despre structura primară a celor trei rămași a completat secvențierea întregului genom.
Din 125 de milioane de perechi de baze, a fost determinată structura primară de 119 milioane, ceea ce reprezintă 92% din întregul genom. Doar 8% din genomul Arabidopsis care conține blocuri mari de segmente repetitive de ADN s-a dovedit a fi inaccesibil pentru studiu. În ceea ce privește completitatea și minuțiozitatea secvențierii genomului eucariotic, Arabidopsis rămâne încă în primii trei campioni împreună cu un organism de drojdie unicelulară. Saccharomyces cerevisiaeȘi organism pluricelular animal Caenorhabditis eleganta(Vezi tabelul).
Aproximativ 15.000 de gene individuale care codifică proteine au fost găsite în genomul Arabidopsis. Aproximativ 12.000 dintre acestea sunt conținute sub formă de două copii per genom haploid (unic), astfel încât numărul total genele este de 27 mii. Numărul de gene din Arabidopsis nu diferă mult de numărul de gene din organisme precum oamenii și șoarecii, dar dimensiunea genomului său este de 25-30 de ori mai mică. Această împrejurare este asociată cu caracteristici importante în structura genelor individuale ale Arabidopsis și structura de ansamblu genomul lui.
Genele Arabidopsis sunt compacte, conținând doar câțiva exoni (regiuni care codifică proteine) separați de segmente scurte (aproximativ 250 bp) de ADN necodificatori (introni). Intervalele dintre genele individuale sunt în medie de 4600 de perechi de baze. Pentru comparație, subliniem că genele umane conțin multe zeci și chiar sute de exoni și introni, iar regiunile intergenice au dimensiuni de 10 mii de perechi de baze sau mai mult. Se presupune că prezența unui genom compact mic a contribuit la stabilitatea evolutivă a Arabidopsis, deoarece ADN-ul său a devenit o țintă pentru diferiți agenți dăunători într-o măsură mai mică, în special pentru introducerea de fragmente de ADN repetitive asemănătoare virusului (transpozoni). în genom.
Printre alte caracteristici moleculare ale genomului Arabidopsis, trebuie remarcat faptul că exonii sunt îmbogățiți în guanină și citozină (44% în exoni și 32% în introni) în comparație cu genele animale, precum și prezența genelor dublu repetate (duplicate). Se presupune că o astfel de dublare a avut loc ca urmare a a patru evenimente simultane, constând în dublarea (repetarea) unei părți a genelor Arabidopsis sau fuziunea genomilor înrudite. Aceste evenimente, care au avut loc în urmă cu 100-200 de milioane de ani, sunt o manifestare a tendinței generale de poliploidizare (o creștere multiplă a numărului de genomi dintr-un organism), care este caracteristică genomului plantelor. Cu toate acestea, unele fapte arată că genele duplicate în Arabidopsis nu sunt identice și funcționează diferit, ceea ce poate fi asociat cu mutații în regiunile lor de reglementare.
Orezul a devenit un alt obiect de secvențiere completă a ADN-ului. Genomul acestei plante este, de asemenea, mic (12 cromozomi, dând un total de 420-470 milioane bp), doar de 3,5 ori mai mare decât cel al Arabidopsis. Cu toate acestea, spre deosebire de Arabidopsis, orezul are o importanță economică deosebită, fiind baza alimentației pentru mai mult de jumătate din umanitate, prin urmare, nu numai miliarde de consumatori, ci și o armată de oameni multimilionare implicați activ în procesul foarte laborios al acestuia. cultivarea sunt extrem de interesați să-și îmbunătățească proprietățile.
Unii cercetători au început să studieze genomul orezului încă din anii 1980, dar aceste studii au atins o amploare serioasă abia în anii 1990. În 1991, în Japonia a fost creat un program de descifrare a structurii genomului orezului, reunind eforturile multor grupuri de cercetare. În 1997, pe baza acestui program a fost organizat Proiectul Internațional al Genomului Orezului. Participanții săi au decis să-și concentreze eforturile pe secvențierea uneia dintre subspeciile de orez ( Oriza sativajaponica), în studiul căruia s-au înregistrat deja progrese semnificative până la acel moment. Un stimulent serios și, la figurat vorbind, o vedetă călăuzitoare pentru o astfel de muncă a fost programul „Genom uman”.
În cadrul acestui program a fost testată strategia de diviziune ierarhică „cromozomială” a genomului, pe care participanții consorțiului internațional au folosit-o pentru a descifra genomul orezului. Cu toate acestea, dacă în studiul genomului uman, fracțiile de cromozomi individuali au fost izolate folosind diverse metode, atunci materialul specific pentru cromozomii individuali de orez și regiunile lor individuale a fost obținut prin microdisecție cu laser (decuparea obiectelor microscopice). Pe o lamă de microscop, unde se află cromozomii de orez, sub influența unui fascicul laser, totul este ars, cu excepția cromozomului sau a secțiunilor acestuia programate pentru analiză. Materialul rămas este folosit pentru clonare și secvențiere.
Au fost publicate numeroase rapoarte cu privire la rezultatele secvențierii fragmentelor individuale ale genomului orezului, efectuate cu mare acuratețe și detaliu, caracteristice tehnologiei ierarhice. Se credea că determinarea structurii primare complete a genomului orezului va fi finalizată până la sfârșitul anului 2003 – mijlocul anului 2004, iar rezultatele, împreună cu datele privind structura primară a genomului Arabidopsis, vor fi utilizate pe scară largă în comparație. genomica altor plante.
Cu toate acestea, la începutul anului 2002, două grupuri de cercetare - unul din China, celălalt din Elveția și Statele Unite - au publicat rezultatele unui proiect complet de secvențiere (aproximativă) a genomului orezului, realizat folosind tehnologia de clonare totală. Spre deosebire de studiul în etape (ierarhice), abordarea totală se bazează pe clonarea simultană a întregului ADN genomic într-unul dintre vectorii virali sau bacterieni și obținerea unui număr semnificativ (uriaș pentru genomi medii și mari) de clone individuale care conțin diverse Segmente de ADN. Pe baza analizei acestor secțiuni secvențiate și a suprapunerii secțiunilor terminale identice ale ADN-ului, se formează un contig - un lanț de secvențe ADN unite între ele. Contigul general (total) este structura primară a întregului genom sau cel puțin a unui cromozom individual.
Într-o astfel de prezentare schematică, strategia clonării totale pare simplă. De fapt, întâmpină dificultăți serioase asociate cu necesitatea obținerii unui număr imens de clone (se acceptă în general că genomul sau regiunea sa studiată trebuie acoperite de clone de cel puțin 10 ori), o cantitate imensă de secvențiere și extrem de complexă. munca la clone de andocare care necesită participarea specialiștilor în bioinformatică. Un obstacol serios în calea clonării totale este o varietate de segmente ADN repetitive, al căror număr, după cum sa menționat deja, crește brusc pe măsură ce dimensiunea genomului crește. Prin urmare, strategia de secvențiere totală este utilizată în principal în studiul genomului virusurilor și microorganismelor, deși a fost folosită cu succes pentru a studia genomul unui organism multicelular, Drosophila.
Rezultatele secvențierii totale a acestui genom au fost „suprapuse” unei game uriașe de informații despre structura sa cromozomială, genică și moleculară, obținute pe o perioadă de aproape 100 de ani de studiu a Drosophila. Și totuși, în ceea ce privește gradul de secvențiere, genomul Drosophila (66% din dimensiunea totală a genomului) este semnificativ inferior genomul Arabidopsis (92%), în ciuda dimensiunilor lor destul de apropiate - 180 de milioane și, respectiv, 125 de milioane de perechi de baze. . Prin urmare, recent s-a propus denumirea tehnologiei mixte, care a fost folosită pentru secvențierea genomului Drosophila.
Pentru a secvenționa genomul orezului, grupurile de cercetare menționate mai sus au luat două dintre subspeciile sale, cele mai cultivate în țările asiatice, - Oriza saliva L. ssp indicajȘi Oriza saliva L. sspjaponica. Rezultatele studiilor lor coincid în multe privințe, dar diferă în multe privințe. Astfel, reprezentanții ambelor grupuri au declarat că au ajuns la aproximativ 92-93% din suprapunerea genomului cu contigs. S-a demonstrat că aproximativ 42% din genomul orezului este reprezentat de repetări scurte de ADN constând din 20 de perechi de baze, iar majoritatea elementelor mobile de ADN (transpozoni) sunt localizate în regiuni intergenice. Cu toate acestea, datele privind dimensiunea genomului orezului diferă semnificativ.
Pentru subspecia japoneză, dimensiunea genomului este determinată a fi de 466 milioane de perechi de baze, iar pentru subspecia indiană, 420 milioane. Motivul acestei discrepanțe nu este clar. Poate fi o consecință a diferitelor abordări metodologice în determinarea dimensiunii părții necodificatoare a genomului, adică nu reflectă adevărata stare de lucruri. Dar este posibil să existe o diferență de 15% în dimensiunea genomilor studiati.
A doua discrepanță majoră a fost dezvăluită în numărul de gene găsite: pentru subspecia japoneză, de la 46 022 la 55 615 gene per genom, iar pentru subspecia indiană, de la 32 000 la 50 000. Motivul acestei discrepanțe nu este clar.
Incompletitudinea și inconsecvența informațiilor primite se remarcă în comentariile la articolele publicate. De asemenea, aici se exprimă speranța că lacunele în cunoașterea genomului orezului vor fi eliminate prin compararea datelor de „secvențiere brută” cu rezultatele secvențierii detaliate, ierarhice, efectuate de participanții la Proiectul Internațional de Genom al Orezului.
GENOMIA PLANTELOR COMPARATIVA SI FUNCTIONALA
Datele extinse obținute, dintre care jumătate (rezultatele grupului chinez) sunt disponibile publicului, deschid fără îndoială perspective largi atât pentru studiul genomului orezului, cât și pentru genomica plantelor în general. O comparație a proprietăților genomului Arabidopsis și al orezului a arătat că majoritatea de gene (până la 80%) identificate în genomul Arabidopsis a fost găsită și în genomul orezului, cu toate acestea, pentru aproximativ jumătate din genele găsite în orez, analogi (ortologi) în genomul Arabidopsis nu au fost încă găsiți. În același timp, 98% dintre genele a căror structură primară a fost stabilită pentru alte cereale au fost găsite în genomul orezului.
Discrepanța semnificativă (aproape dublă) dintre numărul de gene din orez și Arabidopsis este uluitoare. În același timp, datele proiectului de decodificare a genomului orezului, obținute prin secvențierea totală, practic nu sunt comparate cu rezultatele ample ale studiului genomului orezului prin metoda clonării și secvențierii ierarhice, adică ceea ce are nu a fost efectuată cu privire la genomul Drosophila. Prin urmare, rămâne neclar dacă diferența în numărul de gene în Arabidopsis și orez reflectă adevărata stare de lucruri sau dacă este explicată prin diferența de abordări metodologice.
Spre deosebire de genomul Arabidopsis, nu sunt date date despre genele gemene din genomul orezului. Este posibil ca cantitatea lor relativă să fie mai mare în orez decât în Arabidopsis. Această posibilitate este susținută indirect de date privind prezența formelor poliploide de orez. Se poate aștepta mai multă claritate cu privire la această problemă după finalizarea Proiectului Internațional de Genom al Orezului și se obține o imagine detaliată a structurii primare a ADN-ului acestui genom. Motive serioase pentru o astfel de speranță sunt oferite de faptul că, după publicarea lucrărilor privind secvențierea brută a genomului orezului, numărul de publicații privind structura acestui genom a crescut brusc, în special, au apărut informații despre secvențierea detaliată. dintre cromozomii 1 și 4 ai săi.
Cunoașterea, cel puțin aproximativ, a numărului de gene din plante este de o importanță fundamentală pentru genomica comparativă a plantelor. Inițial, s-a crezut că, deoarece toate plantele cu flori sunt foarte apropiate una de cealaltă în ceea ce privește caracteristicile lor fenotipice, genomul lor ar trebui să fie, de asemenea, similar. Și dacă studiem genomul Arabidopsis, vom obține informații despre majoritatea genomilor altor plante. O confirmare indirectă a acestei presupuneri sunt rezultatele secvențierii genomului șoarecelui, care este surprinzător de aproape de genomul uman (aproximativ 30 de mii de gene, dintre care doar 1 mie s-au dovedit a fi diferite).
Se poate presupune că motivul diferențelor dintre genomul Arabidopsis și orezul constă în apartenența lor la diferite clase de plante - dicotiledone și monocotiledone. Pentru a clarifica această problemă, este foarte de dorit să se cunoască cel puțin o structură primară brută a unei alte plante monocotiledonate. Cel mai realist candidat ar putea fi porumbul, al cărui genom este aproximativ egal cu genomul uman, dar totuși mult mai mic decât genomul altor cereale. Valoarea nutritivă a porumbului este binecunoscută.
Vastul material obținut ca urmare a secvențialării genomului Arabidopsis și orezului devine treptat baza unui studiu la scară largă al genomului plantelor folosind genomica comparativă. Astfel de studii au o semnificație biologică generală, deoarece fac posibilă stabilirea principiilor principale ale organizării genomului plantei în ansamblu și a cromozomilor lor individuali, identificarea trăsăturilor comune ale structurii genelor și regiunilor lor de reglare și luarea în considerare a acestora. raportul dintre partea activă funcțional (genă) a cromozomului și diferitele regiuni ADN intergenice care nu codifică proteine. Genetica comparativă devine, de asemenea, din ce în ce mai importantă pentru dezvoltarea genomicii funcționale umane. Pentru studii comparative s-a efectuat secvențierea genomilor de pește-papă și șoarece.
La fel de important este studiul genelor individuale responsabile de sinteza proteinelor individuale care determină funcțiile specifice ale corpului. Este în descoperirea, izolarea, secvențierea și determinarea funcției genelor individuale în care se află semnificația practică, în primul rând medicală, a programului Genomului uman. Această împrejurare a fost remarcată în urmă cu câțiva ani de către J. Watson, care a subliniat că programul Genomului uman va fi finalizat numai atunci când vor fi determinate funcțiile tuturor genelor umane.
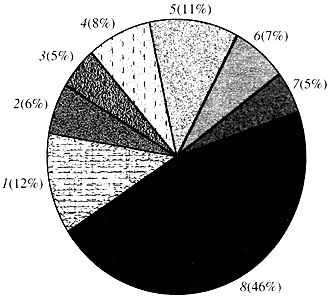
Orez. 4. Clasificare în funcție de funcția genelor Arabidopsis
1 - gene pentru creștere, diviziune și sinteza ADN; 2 - gene de sinteză a ARN (transcripție); 3 - gene pentru sinteza si modificarea proteinelor; 4 - gene pentru dezvoltare, îmbătrânire și moarte celulară; 5 - gene ale metabolismului celular și ale metabolismului energetic; 6 - gene de interacțiune intercelulară și transmitere a semnalului; 7 - gene pentru furnizarea altor procese celulare; 8 - gene cu funcție necunoscutăÎn ceea ce privește funcția genelor plantelor, știm mai puțin de o zecime din ceea ce știm despre genele umane. Chiar și în Arabidopsis, al cărui genom este mult mai studiat decât genomul uman, funcția a aproape jumătate din genele sale rămâne necunoscută (Fig. 4). Între timp, pe lângă genele comune animalelor, plantele au un număr semnificativ de gene care sunt specifice doar (sau cel puțin predominant) pentru acestea. Vorbim despre genele implicate în transportul apei și sinteza peretelui celular, care este absent la animale, despre genele care asigură formarea și funcționarea cloroplastelor, fotosinteza, fixarea azotului, precum și sinteza a numeroase produse aromatice. Această listă poate fi continuată, dar este deja clar cu ce sarcină dificilă se confruntă genomica funcțională a plantelor.
Secvențierea întregului genom oferă informații aproape adevărate despre numărul total de gene dintr-un organism dat, face posibilă plasarea unor informații mai mult sau mai puțin detaliate și fiabile despre structura lor în băncile de date și facilitează munca de izolare și studiere a genelor individuale. Cu toate acestea, secvențierea genomului nu înseamnă în niciun caz să stabilească funcția tuturor genelor.
Una dintre cele mai promițătoare abordări ale genomicii funcționale se bazează pe identificarea genelor de lucru care sunt utilizate pentru transcrierea (citirea) ARNm. Această abordare, inclusiv tehnologie moderna microarrays, vă permite să identificați simultan până la zeci de mii de gene funcționale. Recent, folosind această abordare, a început studiul genomului plantelor. Pentru Arabidopsis, a fost posibil să se obțină aproximativ 26 de mii de transcrieri individuale, ceea ce facilitează foarte mult posibilitatea de a determina funcția aproape tuturor genelor sale. La cartofi, a fost posibil să se identifice aproximativ 20.000 de gene de lucru care sunt importante pentru înțelegerea atât a proceselor de creștere și formare a tuberculilor, cât și a proceselor de boală a cartofului. Se presupune că aceste cunoștințe vor crește rezistența unuia dintre cele mai importante produse alimentare la agenți patogeni.
Dezvoltarea logică a genomicii funcționale a fost proteomica. Acest nou domeniu de știință studiază proteomul, care este de obicei înțeles ca un set complet de proteine dintr-o celulă la un anumit moment. Un astfel de set de proteine, care reflectă starea funcțională a genomului, se schimbă tot timpul, în timp ce genomul rămâne neschimbat.
Studiul proteinelor a fost folosit de mult timp pentru a judeca activitatea genomului plantelor. După cum se știe, enzimele prezente în toate plantele diferă în fiecare specie și soiuri în secvența de aminoacizi. Astfel de enzime, cu aceeași funcție, dar cu o secvență diferită de aminoacizi individuali, se numesc izoenzime. Au proprietăți fizico-chimice și imunologice diferite ( masa moleculara, încărcătură), care poate fi detectată prin cromatografie sau electroforeză. De mulți ani, aceste metode au fost folosite cu succes pentru a studia așa-numitul polimorfism genetic, adică diferențele dintre organisme, soiuri, populații, specii, în special grâul și formele înrudite de cereale. Cu toate acestea, în În ultima vreme Datorită dezvoltării rapide a metodelor de analiză ADN, inclusiv secvențierea, studiul polimorfismului proteic a fost înlocuit cu studiul polimorfismului ADN. Cu toate acestea, studiul direct al spectrelor proteinelor de depozitare (prolamine, gliadine etc.), care determină principalele proprietăți nutriționale ale cerealelor, rămâne o metodă importantă și de încredere pentru analiza genetică, selecția și producția de semințe a plantelor agricole.
Cunoașterea genelor, a mecanismelor de exprimare și reglare a acestora este extrem de importantă pentru dezvoltarea biotehnologiei și producerea de plante transgenice. Se știe că succesele impresionante în acest domeniu provoacă o reacție ambiguă din partea comunității de mediu și medicale. Cu toate acestea, există o zonă a biotehnologiei vegetale în care aceste temeri, dacă nu complet nefondate, atunci, în orice caz, par a fi de puțină importanță. Vorbim despre crearea de plante industriale transgenice care nu sunt folosite ca produse alimentare. India a recoltat recent prima recoltă de bumbac transgenic care este rezistent la o serie de boli. Există informații despre introducerea unor gene speciale care codifică proteine pigmentare în genomul bumbacului și producerea de fibre de bumbac care nu necesită vopsire artificială. Alte cultura tehnica, care poate face obiectul unei inginerie genetică eficientă, este inul. Utilizarea sa ca alternativă la bumbac pentru materii prime textile a fost discutată recent. Această problemă este extrem de importantă pentru țara noastră, care și-a pierdut propriile surse de bumbac brut.
PERSPECTIVE DE STUDIAREA GENOMULUI PLANTELOR
Evident, studiile structurale ale genomului plantelor se vor baza pe abordările și metodele genomicei comparative, folosind ca material principal rezultatele descifrării genomului Arabidopsis și orezului. Un rol important în dezvoltarea genomicii comparative a plantelor îl vor juca, fără îndoială, informațiile care mai devreme sau mai târziu vor fi furnizate de secvențierea totală (aproximată) a genomilor altor plante. În acest caz, genomica comparativă a plantelor se va baza pe stabilirea de relații genetice între loci individuali și cromozomi aparținând unor genomi diferiți. Ne vom concentra nu atât pe genomica generală a plantelor, cât pe genomica selectivă a locurilor cromozomiale individuale. De exemplu, s-a demonstrat recent că gena responsabilă pentru vernalizare este localizată la locusul VRn-AI al cromozomului hexaploid de grâu 5A și locusul Hd-6 al cromozomului 3 de orez.
Dezvoltarea acestor studii va fi un impuls puternic pentru identificarea, izolarea și secvențierea multor gene ale plantelor importante din punct de vedere funcțional, în special, genele responsabile de rezistența la boli, rezistența la secetă și adaptabilitatea la diferite condiții de creștere. Din ce în ce mai mult, se va folosi genomica funcțională, bazată pe detectarea în masă (screening) a genelor care funcționează în plante.
Putem prevedea îmbunătățiri suplimentare ale tehnologiilor cromozomiale, în primul rând metoda de microdisecție. Utilizarea sa extinde dramatic posibilitățile cercetării genomice fără a necesita costuri uriașe, cum ar fi, de exemplu, secvențierea totală a genomului. Metoda de localizare pe cromozomii plantelor a genelor individuale cu ajutorul hibridizării va fi răspândită în continuare. in situ.În prezent, utilizarea sa este limitată de numărul mare de secvențe repetitive din genomul plantei și, eventual, de particularitățile organizării structurale a cromozomilor plantelor.
Tehnologiile cromozomiale vor deveni de mare importanță pentru genomica evolutivă a plantelor în viitorul apropiat. Aceste tehnologii relativ ieftine fac posibilă evaluarea rapidă a variabilității intra și interspecifice, studiul genomilor alopoliploizi complexe de grâu tetraploid și hexaploid, triticale; analiza proceselor evolutive la nivel cromozomial; investighează formarea genomilor sintetici și introducerea (introgresiunea) materialului genetic străin; identificarea relațiilor genetice dintre cromozomii individuali ai diferitelor specii.
Studiul cariotipului plantelor folosind metode citogenetice clasice, îmbogățit prin analiză biologică moleculară și tehnologie computerizată, va fi utilizat pentru caracterizarea genomului. Acest lucru este deosebit de important pentru studierea stabilității și variabilității cariotipului la nivelul nu numai al organismelor individuale, ci și al populațiilor, soiurilor și speciilor. În cele din urmă, este dificil de imaginat cum poate fi estimat numărul și spectrele de rearanjamente cromozomiale (aberații, punți) fără utilizarea metodelor de colorare diferențială. Astfel de studii sunt extrem de promițătoare pentru monitorizare mediu inconjuratorîn funcţie de starea genomului plantei.
ÎN Rusia modernă secvențierea directă a genomului plantelor este puțin probabilă. O astfel de muncă, care necesită investiții mari, depășește puterea economiei noastre actuale. Între timp, datele privind structura genomului Arabidopsis și orezului, obținute de știința mondială și disponibile în băncile internaționale de date, sunt suficiente pentru dezvoltarea genomicii plantelor interne. Putem prevedea extinderea studiilor genomului plantelor bazate pe abordări de genomică comparativă pentru a rezolva probleme specifice de ameliorare și producție a culturilor, precum și pentru a studia originea diferitelor specii de plante de mare importanță economică.
Se poate presupune că abordările genomice precum tiparea genetică (analize RELF, RAPD, AFLP etc.), care sunt destul de accesibile pentru bugetul nostru, vor fi utilizate pe scară largă în practica internă de ameliorare și producția de culturi. În paralel cu metodele directe de determinare a polimorfismului ADN, abordări bazate pe studiul polimorfismului proteic, în primul rând proteinele de depozitare a cerealelor, vor fi utilizate pentru rezolvarea problemelor de genetică și ameliorare a plantelor. Tehnologiile cromozomiale vor fi utilizate pe scară largă. Sunt relativ ieftine, dezvoltarea lor necesită investiții destul de moderate. În domeniul studiilor cromozomiale, știința domestică nu este inferioară lumii.
Trebuie subliniat faptul că știința noastră a adus o contribuție semnificativă la formarea și dezvoltarea genomicii plantelor [ , ].
Rolul fundamental l-a jucat N.I. Vavilov (1887-1943).
În biologia moleculară și genomica plantelor, contribuția de pionierat a lui A.N. Belozerski (1905-1972).
În domeniul studiilor cromozomiale, este de remarcat munca remarcabilului genetician S.G. Navashin (1857-1930), care a descoperit primul cromozomi sateliti la plante și a demonstrat că este posibil să se facă distincția între cromozomi individuali în funcție de caracteristicile morfologiei lor.
Un alt clasic stiinta ruseasca G.A. Levitsky (1878-1942) a descris în detaliu cromozomii de secară, grâu, orz, mazăre și sfeclă de zahăr, a introdus termenul „cariotip” în știință și a dezvoltat doctrina acestuia.
Specialiștii moderni, bazându-se pe realizările științei mondiale, pot aduce o contribuție semnificativă la dezvoltarea ulterioară a geneticii și genomicii plantelor.
Autorul își exprimă sincere mulțumiri academicianului Yu.P. Altukhov pentru discuția critică a articolului și sfaturi valoroase.Munca echipei conduse de autorul articolului a fost susținută de Fundația Rusă pentru Cercetare de bază (granturi nr. 99-04-48832; 00-04-49036; 00-04-81086), Federația Rusă a sustine școli științifice(granturi Nr. 00-115-97833 și NSh-1794.2003.4) și Programe Academia RusăȘtiințe „Markeri genetici moleculari și cromozomiali în dezvoltarea metodelor moderne de ameliorare și producție de semințe”.
LITERATURĂ
1. Zelenin A.V., Badaeva E.D., Muravenko O.V. Introducere în genomica plantelor // Biologie moleculara. 2001. V. 35. S. 339-348. 2. Pen E. Bonanza pentru genomica plantelor // Știință. 1998. V. 282. P. 652-654. 3. Genomica plantelor, Proc. Natl. Acad. sci. STATELE UNITE ALE AMERICII. 1998. V. 95. P. 1962-2032. 4. Cartel N.A. si etc. Genetica. Dicţionar enciclopedic. Minsk: Technologia, 1999. 5. Badaeva E.D., Friebe B., Gill B.S. 1996. Diferențierea genomului în Aegilops. 1. Distribuția secvențelor de ADN foarte repetitive pe cromozomii speciilor diploide // Genom. 1996. V. 39. P. 293-306. Istoricul analizei cromozomilor // Biol. membranelor. 2001. T. 18. S. 164-172.
complet definit. Prin urmare, munca de descifrare a genomului nematodului ar trebui să fie recunoscută ca fiind foarte reușită.
Un succes și mai mare este asociat cu decodificarea genomului Drosophila, doar în
De 2 ori mai mic decât ADN-ul uman și de 20 de ori mai mare decât ADN-ul nematodului. În ciuda un grad înalt Studiul genetic al Drosophila, aproximativ 10% din genele sale erau necunoscute până în acel moment. Dar cel mai paradoxal este faptul că Drosophila, mult mai bine organizată decât nematodul, s-a dovedit a avea mai puține gene decât viermele rotunzi microscopic! Este greu de explicat din pozițiile biologice moderne. Mai multe gene decât la Drosophila sunt prezente și în genomul decodificat al unei plante din familia cruciferelor - Arabidopsis, folosită pe scară largă de geneticieni ca obiect experimental clasic.
Dezvoltarea proiectelor genomice a fost însoțită de dezvoltarea intensivă a multor domenii ale științei și tehnologiei. Deci, bioinformatica a primit un impuls puternic pentru dezvoltarea sa. A fost creat un nou aparat matematic pentru stocarea și procesarea unor cantități uriașe de informații; au fost proiectate sisteme de supercalculatoare cu putere fără precedent; au fost scrise mii de programe care permit în câteva minute să se efectueze o analiză comparativă a diferitelor blocuri de informații, să introducă zilnic noi date în baze de date computerizate,
obținute în diferite laboratoare din întreaga lume și adaptează informații noi la cele acumulate anterior. În același timp, au fost dezvoltate sisteme pentru izolarea eficientă a diferitelor elemente ale genomului și secvențierea automată, adică determinarea secvențelor de nucleotide ADN. Pe această bază, au fost proiectați roboți puternici care accelerează semnificativ secvențierea și o fac mai puțin costisitoare.
Dezvoltarea genomicii, la rândul său, a condus la descoperirea unui număr imens de fapte noi. Semnificația multora dintre ele nu a fost încă evaluată în
viitorul. Dar și acum este evident că aceste descoperiri vor conduce la o regândire a multor poziții teoretice privind apariția și evoluția diferite forme viata pe pamant. Acestea vor contribui la o mai bună înțelegere a mecanismelor moleculare care stau la baza funcționării celulelor individuale și a interacțiunilor acestora; descifrarea detaliată a multor cicluri biochimice necunoscute până acum;
analiza legăturii lor cu procesele fiziologice fundamentale.
Astfel, există o tranziție de la genomica structurală la
funcţional, care la rândul său creează premisele pentru
cercetare baze moleculare funcționarea celulei și a organismului în ansamblu.
Informațiile deja acumulate vor face obiectul analizei pe parcursul
următoarele câteva decenii. Dar la fiecare pas următor
direcția de descifrare a structurii genomurilor diferitelor specii, generează noi tehnologii care facilitează procesul de obținere a informațiilor. Asa de,
utilizarea datelor despre structura și funcția genelor speciilor de ființe vii organizate inferioară poate accelera semnificativ căutarea
deplasa destul de laborios metode moleculare caută gene.
Cea mai importantă consecință a descifrării structurii genomului unei anumite specii este capacitatea de a identifica toate genele acesteia și,
respectiv, identificarea și determinarea naturii moleculare a moleculelor de ARN transcris și a tuturor proteinelor sale. Prin analogie cu genomul, s-au născut conceptele de transcriptom, care unește grupul de molecule de ARN format ca urmare a transcripției, și proteom, care include multe proteine codificate de gene. Astfel, genomica creează fundamentul dezvoltării intensive de noi științe – proteomica și transcriptomica. Proteomica se ocupă cu studiul structurii și funcției fiecărei proteine; analiză compozitia proteinelor celule; determinarea bazei moleculare a funcționării unei singure celule, care este
rezultatul muncii coordonate a multor sute de proteine și
studiul formării trăsăturii fenotipice a unui organism,
care este rezultatul muncii coordonate a miliarde de celule.
Foarte important procese biologice apar la nivel de ARN. Analiza lor este subiectul transcriptomicului.
Cele mai mari eforturi ale oamenilor de știință din multe țări ale lumii care lucrează în domeniul genomicii au avut ca scop rezolvarea proiectului internațional „Genom uman”. Progresul semnificativ în acest domeniu este asociat cu implementarea ideii,
propus de J. S. Venter, să caute și să analizeze
secvențe ADN exprimate, care pot fi folosite ulterior ca un fel de „etichete” sau markeri ai anumitor părți ale genomului. O altă abordare independentă și nu mai puțin fructuoasă a fost luată de munca grupului condus de pr.
Collins. Se bazează pe identificarea primară a genelor pentru bolile ereditare umane.
Descifrarea structurii genomului uman a dus la o descoperire senzațională. S-a dovedit că genomul uman conține doar 32.000 de gene, ceea ce este de câteva ori mai mic decât numărul de proteine. În același timp, există doar 24.000 de gene care codifică proteine; produsele genelor rămase sunt molecule de ARN.
Procentul de similaritate în secvențele de nucleotide ADN între diferiți indivizi, grupuri etnice și rase este de 99,9%.
Această asemănare este ceea ce ne face oameni - Homo sapiens! Toată variabilitatea noastră la nivel de nucleotide se încadrează într-o cifră foarte modestă - 0,1%.
Astfel, genetica nu lasă loc ideilor de superioritate națională sau rasială.
Dar, uitați-vă unul la altul - toți suntem diferiți. Diferențele naționale și cu atât mai mult rasiale sunt și mai vizibile. Deci câte mutații determină variabilitatea unei persoane nu în termeni procentuali, ci în termeni absoluti? Pentru a obține această estimare, trebuie să vă amintiți care este dimensiunea genomului. Lungimea unei molecule de ADN uman este
3,2x109 perechi de baze. 0,1% din aceasta reprezintă 3,2 milioane de nucleotide. Dar amintiți-vă că partea de codificare a genomului ocupă mai puțin de 3% din lungimea totală a moleculei de ADN, iar mutațiile din afara acestei regiuni, de cele mai multe ori, nu au niciun efect asupra variabilității fenotipice. Astfel, pentru a obține o estimare integrală a numărului de mutații care afectează fenotipul, trebuie să luați 3% din 3,2 milioane de nucleotide, ceea ce ne va oferi o cifră de ordinul a 100 000. Adică aproximativ 100 de mii de mutații formează fenotipul nostru. variabilitate. Dacă comparăm această cifră cu numărul total de gene, se dovedește că în medie există 3-4 mutații pe genă.
Care sunt aceste mutații? Marea majoritate (cel puțin 70%)
determină variabilitatea noastră individuală non-patologică, ceea ce ne deosebește, dar nu ne înrăutățește unul în raport cu celălalt. Aceasta include caracteristici precum ochi, păr, culoarea pielii, tipul corpului, înălțimea, greutatea,
un tip de comportament care este, de asemenea, în mare măsură determinat genetic și multe altele. Aproximativ 5% dintre mutații sunt asociate cu boli monogenice. Aproximativ un sfert din mutațiile rămase aparțin clasei de polimorfisme funcționale. Ele sunt implicate în formarea predispoziției ereditare la o patologie multifactorială larg răspândită. Desigur, aceste estimări sunt destul de aspre.
dar ne permit să judecăm structura variabilității ereditare umane.
Capitolul 1.16. Fundamentele genetice moleculare ale evoluției
Revoluția în biologia moleculară care a avut loc la începutul mileniului, culminând cu descifrarea structurii genomului a multor sute de specii de microorganisme, precum și a unor tipuri de protozoare,
drojdia, plantele, animalele și oamenii, au transformat multe dintre ideile tradiționale ale geneticii clasice și au adus mai aproape posibilitatea de a studia mecanismele moleculare ale evoluției și speciației. S-a născut o nouă știință - genomica comparativă,
permiţând înregistrarea apariţiei în diverse linii filogenetice a unor evenimente semnificative evolutiv petrecute la nivelul moleculelor individuale. S-a dovedit că, în cazul general, progresul evolutiv este asociat nu numai, și nu atât de mult cu o creștere a numărului, lungimii și chiar complexității organizării structurale a genelor, ci într-o măsură mult mai mare cu o schimbare a reglementarea muncii lor, care determină coordonarea și expresia specifică țesutului a zeci de mii de gene. În cele din urmă, acest lucru a condus la apariția în organismele superioare a unor complexe mai complexe, foarte specifice, multifuncționale de proteine care interacționează capabile să îndeplinească sarcini fundamental noi.
Să luăm în considerare natura modificărilor care au loc în procesul de evoluție la trei niveluri informaționale: ADN - ARN - proteină sau genom - transcriptom - proteom. În general, putem spune că pe măsură ce crește complexitatea organizării vieții, crește dimensiunea genomului. Astfel, dimensiunea ADN-ului procariot nu depășește 8x106 bp, devine de două ori mai mare la drojdii și protozoare, de 10-15 ori mai mare la insecte, iar la mamifere creșterea ajunge la 3 ordine de mărime, adică de o mie de ori ( 103).
Cu toate acestea, această relație nu este liniară. Deci, în cadrul mamiferelor, nu mai vedem o creștere semnificativă a dimensiunii genomului. În plus, nu este întotdeauna posibil să se observe relația dintre dimensiunea genomului și complexitatea organizării vieții. Astfel, la unele plante, dimensiunea genomului este cu un ordin de mărime sau chiar cu două ordine de mărime mai mare decât la om. Amintiți-vă că creșterea dimensiunii genomului eucariot în comparație cu procariote are loc în principal datorită apariției secvențelor necodante, adică a elementelor opționale. Am spus deja că în genomul uman, exonii în total nu depășesc 1-3%. Și asta înseamnă că numărul de gene în organismele superioare poate fi doar de câteva ori mai mare decât în microorganisme.
Creșterea complexității organizării eucariote se datorează parțial apariției unui sistem de reglementare suplimentar necesar pentru
asigurarea expresiei genelor specifice țesutului. Una dintre consecințele organizării discontinue a genelor care a apărut la eucariote a fost utilizarea pe scară largă a splicing-ului alternativ și a transcripției alternative. Acest lucru a condus la apariția unei noi proprietăți într-un număr mare de gene - capacitatea de a codifica mai multe izoforme de proteine funcțional diferite. Astfel, cantitatea totală de proteine
adică de mărimea proteomului, cei mai mari pot avea de câteva ori mai multe gene.
La procariote, variabilitatea intraspecifică a numărului de gene este acceptabilă și
diferențe similare între diferite tulpini ale multor microorganisme, în
inclusiv agenții patogeni, poate fi de zeci de procente. În același timp, complexitatea organizării diferitelor tipuri de microorganisme se corelează direct cu numărul și lungimea secvențelor de codificare.
Astfel, variabilitatea fenotipică intra și interspecie este în asociere strictă cu dimensiunile transcriptomului și proteomului, care sunt foarte asemănătoare ca valori. La eucariote, numărul de gene este o trăsătură de specie determinată în mod rigid, iar creșterea complexității evolutive se bazează pe un principiu diferit - utilizarea diferențială pe mai multe niveluri a diferitelor componente ale unui proteom limitat și destul de stabil.
Secvențierea genomilor de nematod și Drosophila a arătat că dimensiunile proteomului la aceste specii foarte diferite sunt foarte apropiate și de doar de două ori mai mari decât la drojdii și unele specii bacteriene. Această regularitate – o creștere semnificativă a complexității organizării diferitelor forme de viață, menținând sau crește relativ ușor dimensiunea proteomului – este caracteristică tuturor evoluțiilor ulterioare până la om. Asa de,
proteoamele umane și cele de șoarece practic nu diferă între ele și sunt de mai puțin de 2 ori mai mari decât proteoamele viermelui microscopic nematod sau muștei fructelor Drosophila. Mai mult, identitatea secvențelor de nucleotide ale ADN-ului uman și
marile maimuțe africane este de 98,5%, iar în zonele de codificare ajunge la 99%. Aceste cifre diferă puțin de valoarea de 99,9%,
determinarea asemănării intraspecifice în secvențele de nucleotide ADN între diferiți indivizi, popoare și rase care locuiesc pe planeta noastră. Deci, care sunt schimbările cheie care reprezintă nu mai mult de 1,5% din întregul genom pentru formarea unei persoane? Răspunsul la această întrebare, aparent, ar trebui căutat nu numai la nivel genomic și proteomic.
Într-adevăr, împreună cu stabilitatea relativă a proteomului, în
În cursul evoluției, există o creștere bruscă a dimensiunii și complexității organizării transcriptomului eucariotic datorită apariției în genom a unei cantități uriașe de ADN transcris și necodificant, precum și a unei extinderi semnificative a clasa de gene care codifică ARN. ARN care nu codifică proteine, a căror sursă principală sunt intronii,
alcătuiesc marea majoritate a transcriptomului organismelor superioare,
ajungând la 97-98% din toate unitățile de transcripție. În prezent, funcțiile acestor molecule sunt analizate intens.
Astfel, schimbările evolutive cheie apar pe fondul unei creșteri a dimensiunii genomului, al unui proteom destul de stabil și al unei creșteri accentuate a dimensiunii transcriptomului (Fig. 31.
Figura 31. Schimbări evolutive care au loc pe trei
niveluri informaţionale În acelaşi timp, trecerea de la formele simple de viaţă la cele mai complexe este evident
se corelează cu apariția și distribuția largă în genom a două achiziții evolutive fundamentale și într-o oarecare măsură interdependente: ADN-ul necodant și elemente repetitive. O consecință directă a acestor modificări care apar la nivel genomic este apariția în procesul de evoluție a unui număr imens de ARN care nu codifică proteine.
Care este baza structurală a acestor transformări evolutive?
Toate tranzițiile evolutive majore: de la procariote la eucariote, de la protozoare la organisme multicelulare, de la primele animale la bilaterale și de la cordate primitive la vertebrate, au fost însoțite de o creștere bruscă a complexității genomului. Aparent, astfel de salturi de evoluție sunt rezultatul unor cazuri rare de fuziune cu succes a genomurilor întregi ale diferitelor specii aparținând unor clase sistematice care s-au separat la o distanță considerabilă unele de altele. Astfel, simbioza dintre Archaea și Bacteria a marcat începutul tranziției de la procariote la eucariote. Evident, mitocondriile, cloroplastele și unele alte organele celulare au apărut și ele ca urmare a endosimbiozei. Proprietatea fundamentală a eucariotelor superioare, diploidia, a apărut dintr-o duplicare genomică bine reglată care a avut loc acum aproximativ 500 de milioane de ani.
Dublările genomice în cadrul unei specii au avut loc destul de des și
exemple în acest sens sunt numeroasele cazuri de poliploidie la plante,
ciuperci și uneori chiar animale. Cu toate acestea, mecanisme potențiale
care conduc la apariția în procesul de evoluție a unor forme fundamental noi de viață nu sunt autopoliploidia, ci hibridizarea și transferul orizontal sau fuziunea genomilor. Este de remarcat faptul că cele mai semnificative transformări evolutive, însoțite de fuziunea genomurilor întregi, au loc în condiții extraordinare, în perioadele de tranziții geologice majore, precum modificări ale concentrației de oxigen atmosferic, glaciarea Pământului sau explozia cambriană.
În condiții geologice relativ calme, dublările genelor individuale sau ale segmentelor cromozomiale cu divergența lor ulterioară se dovedesc a fi mai semnificative pentru evoluție. Comparația secvențelor de nucleotide ale genomurilor secvențiate arată că frecvența dublărilor genelor este destul de mare și, în medie, este de 0,01 per genă pe milion de ani. Marea majoritate a acestora nu se manifestă în următoarele câteva milioane de ani și doar în cazuri rare
cazuri, genele duplicate pot dobândi noi funcții adaptative. Cu toate acestea, o clasă mare de dublări de gene „tăcute” servește ca un fel de fond de rezervă pentru nașterea de noi gene și formarea de noi specii. Genomul uman conține de la 10.000 la 20.000 de copii ale genelor procesate care au apărut prin retropunerea ARNm.
Majoritatea lor aparțin clasei pseudogenelor, adică nu sunt exprimate nici din cauza prezenței mutațiilor, nici datorită inserției în regiuni inactive din punct de vedere transcripțional ale genomului. Cu toate acestea, unele dintre aceste gene sunt active, iar natura exprimării lor și chiar funcțiile pot fi diferite,
decât întemeierea genelor.
Un rol deosebit în evoluția primatelor și a oamenilor îl joacă dublări segmentare aparținând clasei de repetări cu copii reduse (LCR) și
a apărut cu mai puțin de 35 de milioane de ani în urmă. Aceste secvențe sunt blocuri foarte identice de ADN, variind în dimensiune de la una la câteva sute de kilobaze. Cel mai adesea, dublările segmentare sunt localizate în regiuni pericentromerice sau telomerice. cromozomi diferiți, și în total ocupă aproximativ 5% din genomul uman.
Nu s-au găsit dublări segmentare în alți genomi secvenționați.
Cea mai mică unitate de duplicare segmentară, numită duplicon, conține fragmente de gene neprocesate neînrudite și
aceasta o deosebește de alte tipuri cunoscute de secvențe repetate. În anumite condiții, dupliconii pot servi ca surse pentru crearea de noi gene himerice transcrise sau familii de gene din diverse combinații de exoni codificatori prezenți în ei. Potrivit unor estimări, între 150 și 350 de gene pot face distincția între genomul cimpanzeului și cel uman.
Fără a sublinia importanța pentru speciarea faptelor apariției unor noi și dispariției secvențelor de codare vechi, trebuie subliniată posibilitatea reală a existenței altor mecanisme,
jucând un rol decisiv în evoluția eucariotelor.
Unul dintre mecanismele motrice ale evoluției sunt elementele mobile întâlnite la toate speciile studiate în acest sens.
Modificările genomului care însoțesc procesul de speciație pot include reorganizări extinse de cariotip, rearanjamente cromozomiale locale, dublări ale familiilor de gene, modificări ale genelor individuale,
însoțite de nașterea sau pierderea lor, precum și de diferențe în expresia genelor, reglementate atât la nivel de transcripție, cât și la nivel de splicing sau translație. Elementele mobile sunt direct legate de toate aceste procese.
În unele cazuri, elementele transposabile în sine poartă secvențe care codifică enzime a căror prezență este necesară pentru a efectua transpunerea ADN-ului sau retropunerea ARN-ului.
Secvențe similare sunt prezente în genomul retrovirusurilor, LTR-
elemente şi transpozoni. Cea mai numeroasă clasă de elemente transpozabile, Alu-repeats, aparține, de asemenea, grupului de retrotranspozoni. Pentru prima dată Alu-
repetele apar la primate cu aproximativ 50-60 de milioane de ani în urmă dintr-o genă mică care codifică ARN. În procesul de evoluție ulterioară, apar divergențe și amplificare puternică a acestei familii. Trecerea de la primate la oameni este însoțită de o creștere explozivă a numărului
Alu-repeats, al căror număr de copii, potrivit unor estimări, ajunge
1,1 milioane. Repetările Alu ocupă aproximativ 10% din genomul uman, dar distribuția lor este neuniformă, deoarece sunt mai mult asociate cu genele. Aceste elemente sunt rareori prezente în exonii de codificare și se găsesc adesea în introni și regiunile necodante ale ARNm pentru a afecta stabilitatea acestor molecule și/sau eficiența translației. Prezența secvențelor Alu în regiunile de intron ale genelor poate fi însoțită de o schimbare a naturii prelucrării preARN, deoarece aceste secvențe conțin regiuni omoloage situsurilor de splicing donor și acceptor. Inserarea elementelor Alu în regiunile de reglare ale genei poate perturba transcripția, rezultând în
© M.D. Golubovsky
Modificări ereditare non-canonice
M.D. Golubovski
Mihail Davidovici Golubovski, doctor Științe biologice, Cercetător principal
Filiala din Sankt Petersburg a Institutului de Istorie a Științelor Naturale și Tehnologiei Academiei Ruse de Științe.
Genetica ca știință a luat formă acum 100 de ani, după a doua descoperire a legilor lui Mendel. Dezvoltarea sa rapidă a fost marcată în ultimii ani de descifrarea compoziției nucleotidice a ADN-ului genomului a multor zeci de specii. Au apărut noi ramuri de cunoaștere - genomica, paleogenetica moleculară. La începutul anului 2001, ca parte a unui costisitor de 10 ani program international a anunțat decodificarea fundamentală a genomului uman. Aceste realizări, probabil, pot fi comparate cu plimbarea în spațiu a unui om și aterizarea pe Lună.
Ingineria genetică și biotehnologia au schimbat foarte mult fața științei. Iată un episod curios, deja inclus în cel mai recent rezumat: „După 1998, a început o cursă fără precedent între cei 1.100 de oameni de știință din comunitatea globală Human Genome Project și firma de capital privat Celera Genomics”. Firma spera să fie prima care să treacă linia de sosire și să beneficieze de brevetarea fragmentelor de ADN uman. Dar până acum principiul a câștigat: „Ceea ce este creat de natură și Dumnezeu nu poate fi brevetat de om.”
Ar putea Gregor Mendel să-și imagineze un tablou atât de fantasmagoric în care își petrecea încet experimentele an de an în liniștea grădinii mănăstirii? În ce măsură transformă ea auto-dezvoltarea naturală a științei? O analiză totală a ADN-ului genomurilor îndepărtează cu adevărat toate acoperirile? Speranțe că Pinocchio a găsit deja prețuita cheie de aur a ușii secrete, în fața unei realități neprevăzute și a paradoxurilor. La om, doar 3% din ADN-ul genomului codifică proteine și poate încă 20-25% sunt implicați în reglarea acțiunii genelor. Care este funcția și restul ADN-ului o are? Genele din genom sunt uneori comparate cu insule mici dintr-o mare de secvențe inactive și posibil nedorite. Cursa ADN seamănă uneori cu zicala: „adu asta, nu știu ce”.
Obiecțiile scepticilor nu au fost în niciun caz înlăturate. Într-adevăr, cu secvențierea totală, nominalizarea (voi folosi un termen la modă) a unui anumit segment de ADN în „rangul genelor” se realizează numai pe baza unor criterii pur formale (semnele de punctuație genetice necesare transcripției). Rolul, timpul și locul de acțiune al majorității „genelor nominalizate” sunt încă complet neclare.
Dar mai este o problemă. Genomul trebuie înțeles ca întregul sistem ereditar, incluzând nu numai structura unui anumit set de elemente ADN, ci și natura conexiunilor dintre ele, care determină cursul ontogenezei în condiții specifice de mediu. Există o triadă sistemică: elemente, conexiuni între ele și proprietăți de integritate. De aici rezultă o concluzie importantă: cunoașterea structurii genelor la nivel de ADN este necesară, dar deloc suficientă pentru a descrie genomul. Suntem doar în pragul înțelegerii modului dinamic de organizare și a formelor necanonice de moștenire [ , ].
În mod neașteptat la sfârșitul secolului al XX-lea. întrebarea care sunt granițele și spectrul variabilității ereditare a depășit sfera discuțiilor pur academice. Mai întâi în Anglia, apoi în Germania, vitele au trebuit să fie sacrificate din cauza unei anomalii neurodegenerative care putea fi transmisă oamenilor cu carnea animalelor bolnave. Agentul infecțios s-a dovedit a fi nu ADN sau ARN, ci proteine numite prioni (din engleză prioni - proteine particule infecțioase - particule infecțioase cu proteine).
Pentru prima dată, cercetătorii au întâlnit manifestarea lor neobișnuită în anii 60. Dar apoi au încercat să interpreteze acest fenomen în cadrul conceptelor clasice, crezând că acestea sunt „infecții virale lente” ale animalelor sau un tip special de mutații supresoare în drojdie. Acum se dovedește „Fenomenul prion nu este o caracteristică exotică a mamiferelor, ci mai degrabă un caz special al unui mecanism biologic general” moștenire dinamică. Probabil că dogma centrală a geneticii moleculare va trebui completată, ținând cont de posibilitatea transmiterii intra și interspecifice după tipul de infecție.
La începutul anilor ’80, clasicul biologiei moleculare și geneticii, R.B. Khesin, a identificat trei forme de variabilitate ereditară non-canonică: modificări ordonate non-aleatoare ale locilor și regiunilor cromozomilor constând din repetarea ADN-ului; modificarea și moștenirea proprietăților citoplasmei; moștenirea epigenetică a modificărilor locale și generale în ambalarea cromatinei. Apoi au fost adăugate gene mobile, al căror comportament a condus la problema inconsecvenței genomului.
Scopul acestui articol este să arate că forme diferite moștenirea non-mendeliană nu este o excepție, ci o consecință a mai multor idei generale despre organizarea genomului. Modificările ereditare nu se limitează în niciun caz la mutații.
Andre Lvov și rolul descoperirii sale
Printr-o coincidenta uimitoare, in acelasi 1953, au aparut doua articole care au determinat chipul genetica modernă: descoperirea dublei helix ADN de către J. Watson și F. Crick și conceptul de profație și lizogenie a bacteriilor de către A. Lvov (1902-1994), care, în opinia mea, nu este acum mai puțin important pentru biologie, medicina si genetica decat dublu helix ADN.
Lvov a stabilit că un fag poate fi integrat în cromozomul unei bacterii și transmis de-a lungul mai multor generații ca o genă bacteriană normală. În această stare, doar gena represoare funcționează în fag, care blochează activitatea tuturor celorlalți loci ai săi. O bacterie care a inclus un fag în genomul său se numește bacterie lizogenă, iar un fag încorporat se numește profag. O astfel de bacterie lizogenă este protejată de infecția cu alți fagi. Sub influența radiațiilor ultraviolete sau a modificărilor din mediul intern al celulei, represorul este inactivat, blocarea este îndepărtată, iar fagul se înmulțește, provocând moartea celulei. Acum este chiar greu de imaginat cât de revoluționară a fost această descoperire.
Andre Lvov - originar din Rusia, părinții săi au emigrat în Franța la sfârșitul secolului al XIX-lea. Imaginea mamei omului de știință Maria Siminovici este imprimată pentru totdeauna pe pânza artistului V. Serov „Fata luminată de soare” (1888). Maria Yakovlevna Lvova-Siminovici a trăit până la 90 de ani. Cu câteva săptămâni înainte de al Doilea Război Mondial, ea a donat Galeriei Tretiakov scrisori și desene de V. Serov. Tatăl lui Lvov îl cunoștea pe Mechnikov și și-a luat fiul să-l vadă la Institutul Pasteur. Astfel, prin secole și țări, firele culturii se întind și se împletesc. Pentru a mea viata lunga A. Lvov a lucrat succesiv ca protozoolog, bacteriolog, biochimist, genetician și, în final, ca virolog. La Institutul Pasteur, el l-a patronat atât pe J. Monod, cât și pe F. Jacob, care a împărțit Premiul Nobel din 1965 cu maestrul pentru descoperirea operonului.
Începând cu anii 1920, se cunosc tulpini bacteriene care se presupune că poartă fagi în stare latentă și provoacă din când în când liza celulară. Cu toate acestea, descoperitorul bacteriofagului F.D. „Errel a privit fagul doar ca pe un agent letal pentru celulă, nepermițând gândul la starea sa latentă. Această opinie a fost împărtășită la început de clasicul geneticii moleculare M. Delbrück. Faptul este că el și colegii săi din SUA au lucrat cu așa-numiții fagi T, care nu se pot integra în cromozomul bacterian.Datorită „demonului autorității”, lizogenia nu a fost studiată cu scrupulozitate din anii 1920. Pionierul această lucrare, un microbiolog strălucit de la Institutul Pasteur, Eugene Wolman a fost capturat de germani ca evreu în timpul ocupației Parisului și a murit.
După război, Lvov a reluat cercetările asupra purtătorului latent de fagi la Institutul Pasteur. În 1953, el a creat conceptul coerent al profagului, realizând imediat semnificația acestuia pentru teoria virală a cancerului și o serie de patologii virale la om. Schema lui clară a fenomenului de lizogenie este încă prezentată în toate rezumatele geneticii moleculare.
În 1958, F. Jacob și Elias Wolman (fiul lui Eugene Wolman) au introdus termenul de epizom pentru elemente care pot exista fie în stare liberă, fie integrate în genomul gazdei. Ei s-au referit la epizomi ca fagi temperați, factorul sexual al bacteriilor, factori de colicinogenitate, cu ajutorul cărora unele tulpini bacteriene ucid alte bacterii. În cartea remarcabilă „Gender and Genetics of Bacteria”, scrisă în 1961 (și publicată în traducere rusă de binecunoscutul genetician S.I. Alikhanyan chiar anul următor), autorii au prevăzut existența unor elemente asemănătoare epizomilor în organismele superioare, în mod prescient. indicând „elementele de control”, descoperite de B. McClintock la începutul anilor '50 ( Premiul Nobelîn Fiziologie sau Medicină 1983). Cu toate acestea, la vremea aceea nu și-au dat seama cât de adâncă este această analogie. După descoperirea la începutul anilor 1970 a mutațiilor de inserție cauzate de încorporarea ADN-ului viral în genomul celular al bacteriilor, a devenit posibilă construirea unei serii evolutive de tranziții bilaterale: segmente de inserție ale „transpozonilor” ale „plasmidelor” fagilor.
Serii similare de reîncarnări au fost găsite printre eucariote. La Drosophila, elementele mobile ale familiei țiganilor („țigani”) pot exista ca copii încorporate în cromozom; să fie sub forma plasmidelor lor circulare sau liniare complete sau reduse în citoplasmă; în cele din urmă, în cazul mutațiilor individuale „permisive” în genomul gazdei, aceștia sunt capabili să pună o coajă, să devină adevărate retrovirusuri infecțioase și să infecteze gazde străine prin intermediul alimentelor. Asemănarea transpozonilor P din Drosophila și retrovirusul endogen HIV la om (Tabel) face posibilă prezicerea posibilelor evenimente genetice evolutive la populațiile umane, soarta inevitabilului său acum și viitoarele contacte cu genomul străin.
Principiul facultativ și conceptul generalizat al genomului
Multe fapte de variabilitate asociate cu elementele transposabile nu se încadrează în conceptul de mutații ca modificări localizate în structura, numărul sau locația locilor genei. Pentru a combina datele geneticii clasice și „mobile”, în 1985 am propus o clasificare naturală a elementelor genomului, incluzând două subsisteme: elemente obligatorii (genele și regiunile lor reglatoare în cromozomi) și elemente facultative (purtători de ADN și ARN, numărul iar topografia căruia variază în celule sau organisme diferite ale aceleiași specii).
Din această clasificare rezultă consecințe importante, care fac posibilă înțelegerea sau formularea multor fapte neobișnuite din domeniul variabilității ereditare. Să numim câteva dintre ele:
- versatilitatea opționalității. Nu există genomi de specii care constau doar din elemente obligatorii, la fel cum nu există organisme vii care constau doar dintr-un schelet;
- non-identitatea genetică a celulelor fiice. Din întâmplare, ele diferă prin numărul și compoziția elementelor facultative citoplasmatice. Raportul dintre fracțiile elementelor ADN obligatorii și facultative este o trăsătură relativ stabilă a speciei. Având un număr similar de loci genici, speciile înrudite pot diferi în cantitatea de ADN de 2-5 sau de mai multe ori, crescând blocurile repetate și modificându-și topografia genomică. Se observă continuu diferite tranziții între părțile obligatorii și facultative ale genomului. Exemplele cele mai evidente sunt mutațiile genetice datorate introducerii (inserțiilor) elementelor mobile sau înmulțirii (amplificarii) segmentelor cromozomiale și trecerea lor la diferite stări intra și extracromozomiale;
- un tip caracteristic de variabilitate ereditară pentru fiecare dintre cele două subsisteme ale genomului. Mutațiile Morgan sunt ușor de corelat cu componenta obligatorie. Mi-am propus să numesc diverse modificări ereditare ale numărului și topografiei elementelor opționale „variații” (ca în muzică - variații pe o anumită temă). Mutațiile, conform conceptelor clasice, apar, de regulă, întâmplător, cu o frecvență scăzută la indivizii individuali. Natura variațiilor este complet diferită - aici sunt posibile schimbări masive, ordonate, sub influența unei varietăți de factori, inclusiv slabi, non-mutageni (temperatură, regim alimentar etc.);
- natura în două etape a majorității modificărilor ereditare naturale. În primul rând, elementele opționale sunt activate ca fiind cele mai sensibile la schimbările din mediu. Apoi, locii genici încep să fie, de asemenea, afectați indirect. Am ajuns la această concluzie în decursul a mulți ani de observații ale focarelor de mutații în natură. Cele mai multe dintre ele s-au dovedit a fi instabile și au fost cauzate de inserții de elemente mobile care sunt activate în mod misterios din când în când în natură. La Drosophila, aproximativ 70% dintre mutațiile care au apărut spontan în natură sau în laborator sunt asociate cu mișcarea elementelor mobile.
Ideea generalizată a genomului ca un ansamblu de elemente obligatorii și facultative extinde, de asemenea, conceptul de „transfer orizontal”, care include nu numai integrarea genelor străine în cromozomii nucleului. Se poate vorbi de transfer orizontal chiar și în cazurile în care se creează o asociere stabilă a două sisteme genetice, în care apar noi caracteristici și proprietăți.
Opționalitatea funcțională a genomului
Modificările moștenite apar ca urmare a erorilor în procesele care operează cu material ereditar orice organisme vii - replicare, transcriere, traducere, precum și reparare și recombinare.
Replicarea facultativă înseamnă posibilitatea unei hiper- sau hipo-replicații relativ autonome a segmentelor individuale de ADN, indiferent de replicarea regulată planificată a întregului ADN genomic în timpul diviziunii celulare. Astfel de proprietăți sunt posedate de secțiuni de cromozomi cu repetări, blocuri de heterocromatină. În acest caz, replicarea autonomă duce la înmulțirea numărului de segmente individuale și, de regulă, are un caracter adaptativ.
Natura facultativă a transcripției constă în posibilitatea apariției diferitelor ARNm din același șablon datorită prezenței a mai mult de un promotor și splicing alternativ într-un locus dat. Această situație este normală pentru multe gene.
Ambiguitatea (în terminologia lui S.G. Inge-Vechtomov) a traducerii se manifestă în diferite variante de recunoaștere a aceluiași codon, de exemplu, un codon stop sau un codon pentru includerea unui anumit aminoacid în proteina sintetizată. O astfel de translație depinde de condițiile fiziologice din celulă și de genotip.
Conform teoriei procesului de mutație a lui M.E. Lobashev, apariția unei mutații este asociată cu capacitatea unei celule și a structurilor sale ereditare de a repara daune. Rezultă că apariția unei mutații este precedată de o stare în care dauna este fie complet reversibilă, fie poate fi realizată sub forma unei mutații, înțeleasă ca „reparație neidentică”. La începutul anilor 1970, a devenit clar că stabilitatea ADN-ului într-o celulă nu este o proprietate imanentă a moleculelor de ADN în sine - este menținută de un sistem enzimatic special.
De la mijlocul anilor 1970, rolul evolutiv al „erorilor de recombinare” ca inductor al modificărilor ereditare, mult mai puternic decât erorile de replicare a ADN-ului, a început să devină clar.
La nivel molecular, există trei tipuri de recombinare: generală, specifică locului și replicativă. Pentru prima recombinare, generală, regulată (încrucișarea), repararea include rupturi în lanțul ADN, reticulare și reparare a acestora. Necesită regiuni lungi de omologie ADN. Recombinarea specifică locului se mulțumește cu regiuni de omologie scurte, mai multe baze, care, de exemplu, au ADN-ul fagului l și cromozomul bacterian. În mod similar, are loc includerea elementelor mobile în genom și recombinarea locală somatică în ontogenie între genele imunoglobulinei, creând diversitatea lor uimitoare.
Erorile în recombinarea generală pot fi considerate drept consecințe naturale ale structurii extinse liniar a genelor. Apare o dilemă despre care a scris Khesin: se poate considera că recombinările mitotice sunt un tip special de mutageneză sau, dimpotrivă, unele tipuri de mutații (aberații cromozomiale) sunt rezultatul „erorilor” recombinărilor mitotice.
Dacă mișcările elementelor mobile sau recombinarea regiunilor sunt programate în ontogenie, este dificil de clasificat astfel de modificări ereditare. Transformarea sexuală în drojdie a fost mult timp considerată un eveniment mutațional, dar s-a dovedit că, într-un anumit stadiu al dezvoltării ascosporelor, are o probabilitate mare ca urmare a recombinării specifice locului.
Variațiile genomului ca răspuns la provocările de mediu
În teoria evoluției și în genetică, problema conexiunii dintre modificările ereditare și direcția selecției a fost întotdeauna discutată. Conform ideilor darwiniene și post-darwiniene, schimbările ereditare apar în direcții diferite și abia apoi sunt preluate prin selecție. Deosebit de clară și convingătoare a fost metoda replică inventată la începutul anilor 1950 de către familia Lederberg. Cu ajutorul unei cârpe de catifea, au obținut copii exacte - amprente - ale unei semănături experimentale de bacterii pe o cutie Petri. Apoi, una dintre plăci a fost utilizată pentru selecția rezistenței la fagi și a fost comparată topografia punctelor de apariție a bacteriilor rezistente pe placa cu fag și în martor. Aranjamentul coloniilor rezistente la fagi a fost același în cele două plăci replici. Același rezultat s-a obținut și în analiza mutațiilor pozitive la bacteriile cu defecte în orice metabolit.
Descoperirile din domeniul geneticii mobile au arătat că celula ca sistem integral în cursul selecției își poate rearanja genomul în mod adaptativ. Este capabil să răspundă provocării mediului printr-o căutare genetică activă și să nu aștepte pasiv apariția aleatorie a unei mutații care îi permite să supraviețuiască. Și în experimentele soților Lederberg, celulele nu au avut de ales: fie moarte, fie o mutație adaptativă.
În cazurile în care factorul de selecție nu este letal, sunt posibile rearanjamente graduale ale genomului, legate direct sau indirect de condițiile selecției. Acest lucru a devenit clar odată cu descoperirea, la sfârșitul anilor 1970, a unei creșteri treptate a numărului de loci în care sunt localizate genele de rezistență la un agent selectiv care blochează diviziunea celulară. Se știe că metotrexatul, un inhibitor al diviziunii celulare, este utilizat pe scară largă în medicină pentru a opri creșterea celulelor maligne. Această otravă celulară inactivează enzima dihidrofolat reductază (DHFR), care este controlată de o genă specifică.
Rezistența celulelor Leishmania la otrava citostatică (metotrexat) a crescut treptat, iar proporția segmentelor amplificate cu gena de rezistență a crescut proporțional. Nu numai gena selectată a fost multiplicată, ci și regiunile mari de ADN adiacente acesteia, numite ampliconi. Când rezistența la otravă în Leishmania a crescut de 1000 de ori, segmentele extracromozomiale amplificate au alcătuit 10% din ADN-ul din celulă! Se poate spune că dintr-o genă obligatorie s-a format un grup de elemente facultative. A existat o rearanjare adaptivă a genomului în timpul selecției.
Dacă selecția a continuat suficient de mult, unii dintre ampliconi au fost inserați în cromozomul original, iar după ce selecția a fost oprită, rezistența crescută a persistat.
Odată cu îndepărtarea agentului selectiv din mediu, numărul de ampliconi cu gena de rezistență a scăzut treptat într-un număr de generații, iar rezistența a scăzut simultan. Astfel, a fost modelat fenomenul modificărilor pe termen lung, când schimbările masive cauzate de mediu sunt moștenite, dar se estompează treptat într-un număr de generații.
În timpul selecției repetate, o parte din ampliconii rămași în citoplasmă a asigurat replicarea lor rapidă autonomă, iar rezistența a apărut mult mai repede decât la începutul experimentelor. Cu alte cuvinte, pe baza ampliconilor conservați s-a format un fel de memorie de amplicon celular a selecției trecute.
Dacă comparăm metoda replicilor și cursul selecției pentru rezistență în cazul amplificării, atunci se dovedește că contactul cu factorul selectiv a provocat transformarea genomului, a cărui natură s-a corelat cu intensitatea și direcția de selecție.
Discuție despre mutațiile adaptive
În 1988, un articol al lui J. Cairns și al co-autorilor a apărut în revista Nature despre apariția „mutațiilor direcționate” dependente de selecție în bacteria E. coli. Am luat bacterii purtătoare de mutații în gena lacZ a operonului lactozei, incapabile să descompună lactoza dizaharidă. Cu toate acestea, acești mutanți s-au putut împărți pe un mediu cu glucoză, de unde au fost transferați într-un mediu selectiv cu lactoză după una sau două zile de creștere. După selectarea reversurilor lac+, care, așa cum era de așteptat, au apărut în timpul diviziunilor „glucozei”, celulele care nu se dezvoltă au fost lăsate în condiții de foamete de carbohidrați. Mai întâi, mutanții au murit. Dar după o săptămână sau mai mult, a fost observată o nouă creștere din cauza unui focar de reversiuni în gena lacZ. Ca și cum celulele aflate sub stres sever, fără a se diviza (!), au efectuat o căutare genetică și și-au schimbat adaptativ genomul.
Studiile ulterioare ale lui B. Hall au folosit bacterii cu mutații în gena de utilizare a triptofanului (trp). Aceștia au fost plasați pe un mediu lipsit de triptofan și s-a evaluat frecvența revenirilor la normă, care a crescut tocmai în timpul înfometării cu triptofan. Cu toate acestea, condițiile de foame în sine nu au fost cauza acestui fenomen, deoarece pe mediul cu foamete pentru cisteină, frecvența revenirilor la trp+ nu diferă de normă.
În următoarea serie de experimente, Hall a luat mutanți dublu cu deficit de triptofan purtând ambele mutații în genele trpA și trpB și a plasat din nou bacteriile pe un mediu lipsit de triptofan. Doar indivizii la care reversiunile au avut loc simultan în două gene ale triptofanului ar putea supraviețui. Frecvența de apariție a unor astfel de indivizi a fost de 100 de milioane de ori mai mare decât se aștepta, cu o simplă coincidență probabilistică a mutațiilor în două gene. Hall a preferat să numească acest fenomen „mutații adaptive” și ulterior a arătat că ele apar și în drojdii, adică. la eucariote.
Publicațiile lui Cairns și Hall au stârnit imediat o discuție aprinsă. Rezultatul primei sale runde a fost prezentarea unuia dintre cercetătorii de frunte în domeniul geneticii mobile J. Shapiro. El a discutat pe scurt două idei principale. În primul rând, celula conține complexe biochimice, sau sisteme de „inginerie genetică naturală”, care sunt capabile să remodeleze genomul. Activitatea acestor complexe, ca oricare altul funcția celulară, se poate schimba dramatic în funcție de fiziologia celulei. În al doilea rând, frecvența de apariție a modificărilor ereditare este întotdeauna estimată nu pentru o celulă, ci pentru o populație de celule în care celulele pot face schimb de informații ereditare între ele. În plus, transferul orizontal intercelular cu ajutorul virușilor sau transferul segmentelor de ADN este îmbunătățit în condiții de stres. Potrivit lui Shapiro, aceste două mecanisme explică fenomenul mutații adaptativeși să-l readucă la curentul principal al geneticii moleculare convenționale. Care sunt, în opinia sa, rezultatele discuției? „Am găsit acolo un inginer genetic cu o gamă impresionantă de instrumente moleculare complicate pentru a reorganiza molecula de ADN.” .
În ultimele decenii, la nivel celular s-a deschis un tărâm neprevăzut de complexitate și coordonare, care este mai compatibil cu tehnologia informatică decât cu abordarea mecanizată care a dominat crearea sintezei moderne neo-darwiniene. După Shapiro, pot fi numite cel puțin patru grupuri de descoperiri care au schimbat înțelegerea proceselor biologice celulare.
organizarea genomului. La eucariote, locii genetici sunt aranjați după un principiu modular, reprezentând construcții de module de reglare și codare comune întregului genom. Acest lucru asigură asamblarea rapidă a noilor constructe și reglarea ansamblurilor de gene. Locii sunt organizați în rețele ierarhice, conduse de o genă master switch (ca în cazul reglării sexului sau al dezvoltării ochiului). Mai mult, multe dintre genele subordonate sunt integrate în diferite rețele: ele funcționează în diferite perioade de dezvoltare și afectează multe trăsături ale fenotipului.În condițiile apelului mediului, celula acționează intenționat, ca un computer, când, la pornire, se verifică pas cu pas funcționarea normală a principalelor programe, iar în cazul unei defecțiuni, computerul se oprește. . În general, devine evident, deja la nivelul celulei, că zoologul evoluționist francez neconvențional Paul Grasset are dreptate: „A trăi înseamnă a reacționa, nu a fi o victimă.”capacitățile reparatorii ale celulei. Celulele nu sunt nicidecum victime pasive ale influențelor fizice și chimice aleatorii, deoarece au un sistem de reparații la nivel de replicare, transcriere și traducere.
Mobil elemente geneticeși inginerie genetică naturală. Activitatea sistemului imunitar se bazează pe construirea continuă de noi variante de molecule de imunoglobuline bazate pe acțiunea sistemelor biotehnologice naturale (enzime: nucleaze, ligaze, revers transcriptaze, polimeraze etc.). Aceste sisteme folosesc elemente mobile pentru a crea noi structuri moștenite. În același timp, modificările genetice pot fi masive și ordonate. Reorganizarea genomului este unul dintre principalele procese biologice. Sistemele de inginerie genetică naturală sunt reglementate de sisteme de feedback. Deocamdată sunt inactive, dar în momente cheie sau în perioadele de stres sunt activate.
Prelucrarea informațiilor celulare. Poate una dintre cele mai importante descoperiri din biologia celulară este că celula colectează și analizează continuu informații despre starea sa internă și mediul extern, luând decizii cu privire la creștere, mișcare și diferențiere. Deosebit de indicative sunt mecanismele de control al diviziunii celulare, care stau la baza creșterii și dezvoltării. Procesul de mitoză este universal în organismele superioare și include trei etape succesive: pregătirea pentru diviziune, replicarea cromozomilor și finalizarea diviziunii celulare. Analiza controlului genetic al acestor faze a condus la descoperire puncte singulare, în care celula verifică dacă repararea încălcărilor în structura ADN-ului a avut loc în etapa anterioară sau nu. Dacă erorile nu sunt corectate, etapa ulterioară nu va începe. Când deteriorarea nu poate fi eliminată, este lansat un sistem programat genetic de moarte celulară sau apoptoză.
Modalități de apariție a modificărilor ereditare naturale în mediul sistemului - elemente facultative - elemente obligatorii. Elementele facultative sunt primele care percep factorii de mediu nemutagenici, iar variațiile care apar apoi provoacă mutații. Elementele obligatorii afectează și comportamentul elementelor opționale.
Modificări ereditare non-canonice care apar sub influența selecției pentru citostatice și conduc la amplificarea genelor.
Trăsăturile dobândite sunt moștenite
„Istoria biologiei nu cunoaște un exemplu mai expresiv al unei discuții veche de secole despre o problemă decât o discuție despre moștenirea sau nemoștenirea trăsăturilor dobândite”,- aceste cuvinte sunt la începutul cărții celebrului citolog și istoric al biologiei L. Ya. Blyakher. În istorie, poate, se poate aminti o situație similară cu încercări de a transforma elemente chimice. Alchimiștii credeau în această posibilitate, dar postulatul imuabilității a fost stabilit în chimie elemente chimice. Totuși, acum în fizica nuclearași chimie, cercetarea privind transformarea elementelor și analiza evoluției lor este un lucru comun. Cine a avut dreptate în disputa veche de secole? Putem spune că la nivelul interacțiunilor chimice moleculare nu există o transformare a elementelor, dar la nivel nuclear este regula.
O analogie similară apare cu problema moștenirii trăsăturilor care au apărut în cursul ontogenezei. Dacă schimbările ereditare nou apărute sunt reduse doar la mutații ale genelor și cromozomilor, atunci întrebarea poate fi considerată închisă. Dar dacă pornim de la conceptul generalizat al genomului, inclusiv ideea de ereditate dinamică [ , ], problema trebuie revizuită. Pe lângă mutații, există forme variaționale și epigenetice de variabilitate ereditară asociate nu cu modificări ale textului ADN, ci ale stării genei. Astfel de efecte sunt reversibile și ereditare.
Interesant este că Anuarul Internațional de Genetică, publicat la sfârșitul anului 1991, se deschide cu un articol de O. Landman „Moștenirea trăsăturilor dobândite” . Autorul rezumă faptele obținute cu mult timp în urmă în genetică, arătând că „Moștenirea trăsăturilor dobândite este destul de compatibilă cu conceptul modern de genetică moleculară.” Landman ia în considerare în detaliu aproximativ zece sisteme experimentale în care s-a stabilit moștenirea trăsăturilor dobândite. Patru mecanisme diferite pot duce la aceasta: o modificare a structurilor membranei celulare, sau cortexului, studiat de T. Sonneborn la ciliati; Modificări ale ADN-ului, de ex. modificări transmise clonal în natura metilării ADN-ului local (aceasta include fenomenul de imprimare); modificări epigenetice fără modificări ale ADN-ului; pierderea indusă sau achiziționarea de elemente opționale.
Articolul lui Landman ne face, parcă, martorii unei perioade critice de schimbare a postulatelor în genetică, care părea de neclintit ca o piatră. Autorul cu calm, fără entuziasm și fapte noi uimitoare, combină datele vechi și noi într-un sistem, le oferă o interpretare modernă clară. Se poate formula un principiu general: moștenirea trăsăturilor dobândite este posibilă în cazurile în care o anumită trăsătură fenotipică depinde de numărul sau topografia elementelor facultative.
Voi da două exemple instructive despre Drosophila: primul este asociat cu comportamentul virusului sigma, al doilea - cu elemente mobile responsabile pentru sterilitatea hibridă a femelelor și supramutabilitatea.
Studiul interacțiunii virusului sigma cu genomul Drosophila a început cu mai bine de 60 de ani în urmă. Mai întâi, în 1937, geneticianul francez F. Leritje a descoperit diferențe ereditare ascuțite în diferite linii de muște în ceea ce privește gradul de sensibilitate la dioxid de carbon(CO2). Trăsătura a fost moștenită într-un mod bizar: prin citoplasmă, dar nu numai prin linia maternă, ci uneori prin masculi. Sensibilitatea ar putea fi transmisă și prin injectarea hemolimfei și la diferite tipuri de muște de fructe. În aceste cazuri, trăsătura nu a fost transmisă stabil, dar ca urmare a selecției, moștenirea a devenit stabilă.
Moștenirea non-mendeliană a unei trăsături la Drosophila care depinde de o populație de elemente facultative ale genomului. Semnul sensibilității la CO 2 este cauzat de prezența rhabdovirus sigma în citoplasma muștei. Ca urmare a șocului de temperatură într-un stadiu incipient de dezvoltare a Drosophila, reproducerea virusului este blocată, iar indivizii crescuți dobândesc rezistență la acesta.Sensibilitatea la CO 2 a fost asociată cu reproducerea stabilă în celulele germinale și somatice ale rabdovirusului sigma în formă de glonț care conține ARN, care este similar într-un număr de proprietăți cu virusul rabiei la mamifere. Oogonia (celule din care se formează ouăle în timpul meiozei și maturării) la femelele de linie stabilizată conțin de obicei 10-40 de particule virale, iar ovocite (ouă mature) - 1-10 milioane.Virusul sigma este un element opțional tipic. Mutațiile din genomul său conduc la forme complexe de comportament al sistemului. Au fost găsite cazuri de purtători de virus în care Drosophila rămâne rezistentă la CO2, dar în același timp imună la infecția cu alte tulpini de virus. Situația este destul de comparabilă cu comportamentul sistemului fago-bacterii, care a fost imediat observat de F. Jacob și E. Volman.
Relația dintre genomul Drosophila și virusul care se reproduce în citoplasma sa respectă regulile geneticii intracelulare. Impacturile în timpul ontogenezei pot determina o schimbare a numărului și a topografiei intercelulare a particulelor și, ca urmare, pot modifica gradul de sensibilitate la dioxidul de carbon. Astfel, temperatura ridicată blochează replicarea particulelor virale. Dacă femelele și masculii sunt ținuți la o temperatură de 30°C timp de câteva zile în timpul gametogenezei, descendenții unor astfel de muște vor fi liberi de virus și rezistenți la CO 2 . Așa dobândit în timpul dezvoltarea individuală trăsătura este moștenită de-a lungul unui număr de generații.
Situația cu virusul sigma nu este izolată. Geneticienii francezi au studiat factorii de sterilitate feminină asociați cu comportamentul elementelor mobile de tipul „I”. Moștenirea acestei trăsături este determinată de interacțiuni nuclear-citoplasmatice complexe. Dacă elementele I active sunt localizate în cromozomii paterni, atunci pe fundalul citoplasmei R încep să fie activate, suferă transpoziții multiple și, ca urmare, provoacă perturbări ascuțite în ontogenie la descendenții femelelor cu citoplasmă sensibilă. Astfel de femele depun ouă, dar unii dintre embrioni mor într-un stadiu incipient al clivajului - chiar înainte de formarea blastomerului. Liniile izolate din populațiile naturale diferă în ceea ce privește puterea factorilor I și gradul de reactivitate (sau sensibilitate) a citoplasmei. Acești indicatori pot fi modificați prin influență externă. Vârsta femelelor parentale inițiale, precum și efectul temperaturii ridicate în perioada timpurie de dezvoltare, afectează nu numai fecunditatea femelelor crescute, ci și fecunditatea puilor lor. Cauzate de condițiile de mediu, modificările reactivității citoplasmei sunt menținute pe parcursul multor generații de celule. „Cel mai remarcabil lucru este că aceste modificări în reactivitatea citoplasmei sub influența factorilor negenetici sunt moștenite: se observă moștenirea trăsăturilor „dobândite””,- a notat R.B. Khesin.
Moștenirea prin citoplasmă: de la bunici la nepoți
În teoria dezvoltării și fenogenetică a secolului al XX-lea. un loc important îl ocupă studiile profunde și complet originale ale embriologului P.G.Svetlov (1892-1972). Să ne oprim asupra teoriei cuantizării ontogenezei dezvoltată de el (prezența unor perioade critice în dezvoltare, când are loc determinarea proceselor morfogenetice și în același timp crește sensibilitatea celulelor la agenții nocivi) și asupra ideii dezvoltate în legătură cu acest lucru că studiul ontogenezei ar trebui efectuat nu din momentul fertilizării și formării unui zigot, și, de asemenea, din gametogeneză, inclusiv oogeneză la femelele din generația anterioară - perioada proembrionară.
Pe baza acestor postulate, Svetlov a efectuat experimente simple și clare în anii 1960 pe Drosophila și șoareci. El a arătat în mod convingător că moștenirea persistentă non-mendeliană a proprietăților citoplasmei este posibilă, iar modificările severității trăsăturilor mutante care au apărut după un impact extern pe termen scurt în timpul unei perioade critice de dezvoltare a organismului sunt, de asemenea, transmise într-un număr. de generatii.
Într-unul din seria de experimente, el a comparat gradul de manifestare a trăsăturii mutante la descendenții a două linii de șoareci heterozigoți pentru mutația recesivă a microftalmiei (dimensiunea redusă a retinei și a ochilor din momentul nașterii): fenotip- heterozigoți normali, ale căror mame au fost mutante și cei ale căror mame au fost mutate. Progenitul de la bunica mutantă diferă într-o manifestare mai puternică a trăsăturii. Svetlov a explicat acest fapt ciudat prin faptul că gameții feminini ai femelelor heterozigote se aflau încă în corpul mamelor lor mutante și au fost influențați de ei, ceea ce a crescut mutațiile la nepoții lor.
În esență, Svetlov a stabilit un fenomen care mai târziu a devenit cunoscut sub numele de „imprinting genomic” - o diferență în expresia unei gene, în funcție de faptul că a venit la urmaș de la mamă sau de la tată. Aceste lucrări, din păcate, au rămas subestimate.
Interesant este că încă de la sfârșitul anilor 1980, imprimarea, după cum a remarcat cu inteligență K. Sapienza, un cercetător al acestui fenomen, a fost „Considerată în mod obișnuit o curiozitate genetică care afectează doar câteva trăsături. Am fost întrebat în repetate rânduri de ce pur și simplu îmi pierd timpul cu un fenomen atât de nesemnificativ”. Majoritatea cercetătorilor au acceptat necondiționat una dintre principalele propuneri ale lui Mendel - „rudimentul”, sau gena, nu își poate schimba potența în funcție de sex, pe care se bazează împărțirea observată pe scară largă a 3:1. Dar Sapienza a remarcat pe bună dreptate că atunci când analizează diviziunea mendeliană, de obicei iau în considerare doar prezența sau absența unei trăsături, iar dacă aceasta este cantitativă, atunci granița. da nu setat la pragul acceptat. Dacă, totuși, pentru a dezvălui gradul de manifestare a trăsăturii, se va dezvălui influența amprentei genomice.
Exact aceasta a fost abordarea lui Svetlov când a studiat cu atenție modul în care severitatea trăsăturilor la descendenți se modifică în funcție de genotipul matern. În calitate de embriolog, a văzut comunitatea modificărilor ereditare și speciale neereditare - fenocopii (simularea mutațiilor), dacă este afectat același aparat morfogenetic responsabil de implementarea unei anumite trăsături.
Pentru prima dată la diferite specii de animale (Drosophila și șoareci), Svetlov a arătat posibilitatea moștenirii prin meioză a naturii modificate a manifestării genei mutante. Nu degeaba Khesin a numit aceste lucrări remarcabile în rezumatul său.
Încălzirea pe termen scurt (20 de minute) a corpului unei femele de șoarece în vârstă de opt zile a provocat modificări persistente ale ovocitelor, care au slăbit efectul unei mutații dăunătoare la nepoți! „Transmiterea îmbunătățirii dezvoltării ochilor observată în experimentele cu încălzire poate fi explicată doar prin transmiterea proprietăților dobândite prin moștenire în ovocitele femelelor încălzite”. Svetlov a asociat acest fenomen cu particularitățile formării și structurii oului la animale, deoarece „În ovocit există, parcă, un cadru care reflectă cele mai generale trăsături ale arhitecturii organismului în construcție.” Pentru prevenirea tulburărilor de dezvoltare la om, el a fundamentat necesitatea studierii perioadelor critice ale gametogenezei, în care sensibilitatea la daune este crescută. Poate că, în patogeneza anomaliilor de dezvoltare la om, stadiul formării gameților este chiar mai important decât embriogeneza.

Schema de experimente de P.G. Svetlov, care demonstrează transmiterea unei mutații într-o serie de generații de șoareci - microftalmie. O singură expunere de 20 de minute la temperatură ridicată la șoarecii mutanți de 8 zile are ca rezultat îmbunătățirea dezvoltării ochilor la descendenții lor (F1 și F2). Această trăsătură este moștenită numai prin linia maternă și este asociată cu o modificare a ovocitelor.Astăzi, această concluzie este confirmată de studiile genetice moleculare din ultimul deceniu. Drosophila are trei sisteme de gene materne care formează eterogenitatea axială și polară a citoplasmei și gradienții de distribuție a produselor genetice active biologic. Cu mult înainte de începerea fertilizării, determinarea moleculară (predeterminarea) planului structural și etapele inițiale dezvoltare. În formarea ovocitului, produsele genice ale celulelor organismului matern joacă un rol important. Într-un fel, acest lucru poate fi comparat cu un grup de albine lucrătoare care hrănesc o matcă într-un stup.
La om, celulele germinale primare, din care apar apoi gameții de ou, încep să se separe într-un embrion de două luni. La vârsta de 2,5 luni intră în meioză, dar imediat după naștere, această diviziune este blocată. Se reia dupa 14-15 ani odata cu debutul pubertatii, cand ovulele parasesc foliculii o data pe luna. Dar la sfârșitul celei de-a doua diviziuni, meioza se oprește din nou și blocarea ei este îndepărtată doar atunci când întâlnește spermatozoizi. Astfel, meioza feminină începe la 2,5 luni și se termină abia după 20–30 de ani sau mai mult, imediat după fertilizare.
Zigotul aflat în stadiul de două până la opt celule are o imunitate genomică slăbită. Când am studiat mutațiile de inserție instabile în populațiile naturale de Drosophila, am constatat că activarea unui element mobil, însoțită de o tranziție mutațională, are loc adesea deja în primele diviziuni ale zigotului sau în primele diviziuni ale celulelor germinale primare. Ca rezultat, un eveniment mutant captează imediat o clonă de celule germinale primare, grupul de gameți devine mozaic, iar modificările ereditare ale descendenților apar în ciorchini sau ciorchini, imitând moștenirea familiei.
Aceste experimente sunt foarte importante pentru epidemiologie, atunci când se pune întrebarea cu privire la gradul de influență a unei anumite epidemii virale asupra fondului genetic al descendenților. Studiile de pionier ale S.M. Gershenzon și Yu.N. Aleksandrov, începute la începutul anilor 1960, au condus la concluzia că virusurile care conțin ADN și ARN și acizii lor nucleici sunt agenți mutageni puternici. Intrând în celulă, ele provoacă stres genomic, activează sistemul de elemente mobile ale gazdei și provoacă mutații de inserție instabile într-un grup de loci selectați specifici fiecărui agent.
Acum imaginați-vă că vrem să evaluăm impactul asupra variației genetice umane al unei pandemii virale (de exemplu, gripa). În acest caz, se poate aștepta ca frecvența alt fel anomaliile de dezvoltare vor fi crescute în prima generație la descendenții născuți la un an sau la un an după epidemie. Evaluarea frecvenței modificărilor mutaționale și variaționale în celulele germinale (gameți) ar trebui efectuată la nepoți.

Schema oogenezei în trei generații feminine succesive. P - bunica, F1 - mama, F2 - fiica.
Concluzia generală este că variabilitate ereditară la nepoți pot fi foarte dependente de condițiile în care s-a produs oogeneza la bunicile lor! Imaginează-ți o femeie care în 2000 avea aproximativ 25 de ani și va deveni mamă în mileniul trei. Ovulul fecundat, din care ea însăși s-a născut, a început să se formeze într-o perioadă în care mama ei era încă un embrion de două luni, adică. cândva la mijlocul anilor 1950. Și dacă gripa a făcut furori în acești ani, atunci consecințele ei ar trebui să se simtă într-o generație. Pentru a evalua consecințele epidemiei globale asupra fondului genetic uman, este necesar să se compare nepoții a trei grupuri sau cohorte - cei ale căror bunici au fost însărcinate în anul în care a izbucnit epidemia, cu cei ale căror bunici au rămas însărcinate înainte și după pandemie (acestea sunt două cohorte de control). Din păcate, astfel de date epidemiologice și genetice importante pentru protecția sănătății nu sunt încă disponibile.
Despre fantome și luptă cu monștri
Au trecut treizeci de ani de la experimentele lui Svetlov, care erau simple ca tehnică, dar originale ca concept și profunde în concluziile lor. La mijlocul anilor 1990, a avut loc o cotitură psihologică: numărul lucrărilor din domeniul variabilității ereditare cu cuvântul „epigenetic” în titlu a crescut brusc.
Diferite tipuri de epimutații (variații ereditare ale naturii activității genelor care nu sunt asociate cu modificări ale textului ADN și sunt masive, direcționate și reversibile) s-au mutat de la categoria de fenomen marginal la un fenomen studiat activ. A devenit evident că sistemele vii au „memorie” operațională care este în contact continuu cu mediul și utilizează mijloacele ingineriei embriogenetice naturale pentru o tranziție rapidă ereditară de la un mod de funcționare la altul. Sistemele vii nu sunt victime pasive selecție naturală, și toate formele evolutive de viață nu sunt deloc „Ștergeți pentru o scurtă zi de evadare”, așa cum a scris Mandelstam în celebra sa capodopera Lamarck.
S-a dovedit că epimutațiile pot fi găsite foarte des în „genele clasice” obișnuite, trebuie doar să alegeți un sistem experimental adecvat. În 1906, cu cinci ani înainte ca Morgan să înceapă să lucreze cu Drosophila, biologul evoluționist francez L. Keno a descoperit mutația Mendelian a corpului galben la șoareci. Ea a avut o trăsătură uimitoare - dominanță în raport cu culoarea normală (gri-maro) și letalitatea la homozigot. Când șoarecii galbeni heterozigoți au fost încrucișați între ei, din cauza morții homozigoților, șoarecii normali au apărut la urmași într-un raport nu de 3:1, ci de 2:1. Ulterior, s-a dovedit că multe mutații dominante în diferite organisme se comportă astfel.
S-a dovedit că în regiunea de transcripție a uneia dintre alelele genei „corp galben” a fost introdus un element mobil, care seamănă cu un retrovirus ca structură și proprietăți. Ca urmare a acestei inserții, gena a început să se supună semnelor de punctuație ale intrusului său și s-a activat în mod imprevizibil. „la momentul nepotrivit și la locul nepotrivit.” Mutanții cu inserții dezvoltă multiple defecte (blană galbenă, obezitate, diabet etc.), iar comportamentul lor devine instabil. Activitatea de inserție inutilă este stinsă în diferite grade în diferite țesuturi datorită modificării reversibile sau metilării bazelor ADN. La nivel de fenotip, manifestarea alelei dominante variază foarte mult și este de natură mozaică. Geneticienii australieni au descoperit că femelele galbene selectate dintr-o linie omogenă au avut mai mulți șoareci galbeni la descendenți, iar fenotipul tatălui - purtător al mutației - nu a afectat schimbarea culorii la urmași. Femelele s-au dovedit a fi mai inerțiale, iar ele, selectate în funcție de fenotipul de modificare a ADN-ului, sau amprentele, s-au păstrat mai bine în oogeneză. Alți geneticieni au găsit și o influență pur maternă, similară cu cea găsită în experimentele lui Svetlov. În funcție de dieta femeilor însărcinate, severitatea mutației „corp galben” s-a schimbat într-un anumit fel în genotipul heterozigoților. Această stare alterată este instabilă, dar moștenită la descendenți. Gradul de manifestare a trăsăturii corelat cu gradul de metilare a bazelor ADN din insert.
Referindu-se la acestea și la alte experimente similare, recenzorul științific al revistei „Science” și-a numit articolul „Was Lamarck still a little right?” Această tactică este de înțeles. În primul rând, prudența este justificată atunci când vine vorba de revizuirea a ceea ce a fost considerat ferm stabilit de decenii. În al doilea rând, moștenirea caracteristicilor dobândite este asociată nu numai cu numele lui Lamarck, ci și cu fantoma lui Lysenko (autorul notei îl menționează pe acesta din urmă). Într-adevăr, voluntar sau involuntar, umbra „biologiei Michurin” apare atunci când se discută problema moștenirii trăsăturilor dobândite. Și nu numai în Rusia, unde amintirea tragediei din biologie asociată cu dominația lui Lysenko este încă vie.
Astăzi, multe prevederi general acceptate ale geneticii clasice, pe care Lysenko le-a respins, au devenit involuntar, în ciuda lui, considerate aproape adevăr absolut. Și totuși, dacă unul sau altul cercetător serios a descoperit ceva în exterior în consonanță cu opiniile lui Lysenko, îi era teamă să-l facă public, temându-se de ostracismul din partea comunității științifice. Și chiar dacă lucrarea a fost publicată, a fost însoțită de multe rezerve și a rămas la periferia științei.
Făcând cunoștință în anii 60 cu articolele lui A.A. Lyubishchev (cel mai apropiat prieten al lui Svetlov), am încercat să înțeleg de ce, fiind unul dintre cei mai activi critici autopublicați ai lisenkoismului din 1953 până în 1965, articolele și scrisorile sale au fost adunate în carte. „În apărarea științei” (L., 1990), - cu toate acestea, nu a considerat problema moștenirii trăsăturilor dobândite în cele din urmă rezolvată. Acest expert recunoscut universal în biologie evoluționistă a subliniat incompletitudinea teoriei eredității, asemănarea variabilității ereditare și a modificării. Acum știm cât de dificil este în multe cazuri să tragem o linie între ele. Lyubishchev a citat faptele transformărilor în masă, rapide și ordonate ale fenotipului în evoluție, clar inexplicabile din punctul de vedere al mutațiilor Morgan și al selecției darwiniene. După ce și-a ridicat vocea împotriva monopolului lui Lisenko, Liubishchev a vorbit în apărarea științei ca atare, împotriva regimului Arakcheev care se instalase în ea. În domeniul științei în sine, el a urmat principiul străvechi: „Platon este prietenul meu, dar adevărul este mai drag”.
9. McClintock b.// Știință. 1984. V.226. P.792-801.
10. Cairns J.// Natură. 1988.V.27. P.1-6.
11. Sala D.// Genetica. 1990. V.126. P.5-16
12. Shapiro J.// Știință. 1995. V.268. P.373-374.
12. Blyakher L. Ya. Problema moștenirii trăsăturilor dobândite. M., 1971.
13. Landman O.// Ann. Pr. Genet. 1991. V.25. P.1-20.
14. Sokolova K.B. Dezvoltarea fenogeneticii în prima jumătate a secolului XX. M., 1998.
15. Sapienza K.// În lumea științei. 1990. ?12. pp.14-20.
16. Svetlov P.G.// Genetica. 1966.?5. S.66-82.
17. Korochkin L.I. Introducere în genetica dezvoltării. M., 1999.
) găsit în genomul muștei fructelor ( Drosophila ananassae) o copie completă a genomului bacterian parazit Wolbachia.
Bacteria Wolbachia trăiește în citoplasma celulelor gazdă și este cunoscută pentru că a învățat să regleze fin reproducerea, dezvoltarea și chiar evoluția gazdelor sale. Prin urmare, este adesea numit „microb-manipulator” sau „stăpânul muștelor” (din moment ce trăiește în celule de insecte).
Studiul a început când Julie Dunning-Hotopp, de la JCVI, a descoperit cum anumite gene Wolbachia „cooperează” cu genele Drosophila ca și cum ar fi parte din același genom.
Michael Clark - cercetător la Universitatea din Rochester - a stabilit colonia Drosophila ananassaeîn laborator pentru a afla secretul cu Warren.
Gena Wolbachia din genomul Drosophila (ilustrat de Universitatea din Rochester).
„Timp de câteva luni, am crezut că am greșit cu ceva”, spune Clarke, „am sugerat chiar că s-a dezvoltat rezistență la antibiotice, deoarece am găsit fiecare genă Wolbachia din nou și din nou. Când am luat în sfârșit șervețele pe care le lăsasem singur cu câteva luni în urmă, nu am găsit Wolbachia în sine.
Acum Warren și Clarke încearcă să-și dea seama care este avantajul inserării unei piese atât de mari de ADN pentru Drosophila - poate că genele „străine” oferă gazdei câteva oportunități noi.

Și astfel genele Wolbachia trec în ADN-ul gazdei (ilustrare de Nicolle Rager Fuller, National Science).
Rezultatele studiului au fost publicate într-un articol din revista Science. În ea, autorii sugerează că transferul de gene orizontal (transferul de gene între specii neînrudite) are loc între bacterii și organisme multicelulare din lumea noastră mult mai des decât se credea anterior.
Descifrarea mecanismelor genetice moleculare ale manipulărilor efectuate de Wolbachia cu gazdele sale va oferi omului noi mijloace puternice de influențare a organismelor vii și a naturii în ansamblu.
Cu toate acestea, nu toate insectele sunt sensibile influenta negativa wolbachia. De exemplu, fluturii din Insulele Samoan au „învățat” să-și protejeze masculii. Mă întreb dacă țânțarii de malarie, pe care vor să-i infecteze cu această bacterie, vor învăța să lupte cu ea?
La 50 de ani de la descoperirea structurii ADN-ului
A.V. Zelenin
GENOMUL PLANTELOR
A. V. Zelenin
Zelenin Alexandru Vladimirovici- d.b.n.,
şef al laboratorului Institutului de Biologie Moleculară. V.A. Engelhardt RAS.
Realizările impresionante ale programului „Genom uman”, precum și succesul lucrărilor de descifrare a așa-numitelor genomuri extra-mici (virusuri), mici (bacterii, drojdie) și medii (viermi rotunzi, Drosophila), au făcut posibilă trecerea la un studiu pe scară largă a genomului plantelor mari și foarte mari. Necesitatea urgentă a unui studiu detaliat al genomilor celor mai importante plante din punct de vedere economic a fost subliniată la o întâlnire despre genomica plantelor, organizată în 1997 în Statele Unite [ , ]. De-a lungul anilor care au trecut de atunci, s-au obținut succese neîndoielnice în acest domeniu. În 2000, a apărut o publicație privind secvențierea completă (determinarea secvenței nucleotidice liniare a întregului ADN nuclear) a genomului muștarului mic - Arabidopsis, în 2001 - privind secvențierea preliminară (proiect) a genomului orezului. Au fost raportate în mod repetat lucrări de secvențiere a genomului plantelor mari și super-mari (porumb, secară, grâu), cu toate acestea, aceste rapoarte nu conțineau informații specifice și erau mai degrabă de natura declarațiilor de intenție.
Se presupune că decodificarea genomului plantelor va deschide perspective largi pentru știință și practică. În primul rând, identificarea de noi gene și lanțul de reglare genetică a acestora va crește semnificativ productivitatea plantelor prin utilizarea abordărilor biotehnologice. Odată cu descoperirea, izolarea, reproducerea (clonarea) și secvențierea genelor responsabile pentru funcții atât de importante ale organismului vegetal precum reproducerea și productivitatea, procesele de variabilitate, rezistența la factorii de mediu adversi, precum și împerecherea omoloagă a cromozomilor, apariția se asociază noi oportunităţi de îmbunătăţire a procesului de ameliorare . În cele din urmă, genele izolate și donate pot fi folosite pentru a obține plante transgenice cu proprietăți fundamental noi și pentru a analiza mecanismele de reglare a activității genelor.
Importanța studierii genomului plantelor este subliniată și de faptul că până în prezent numărul de gene de plante localizate, clonate și secvențiate este mic și variază, după diverse estimări, între 800 și 1200. Acesta este de 10-15 ori mai mic decât, pt. exemplu, la oameni.
Statele Unite rămân liderul incontestabil în studiul pe scară largă a genomului plantelor, deși studii intensive ale genomului orezului sunt efectuate în Japonia și, în ultimii ani, în China. La descifrarea genomului Arabidopsis, pe lângă laboratoarele din SUA, au participat activ grupurile de cercetare europene. Conducerea aparentă a Statelor Unite provoacă îngrijorare serioasă a oamenilor de știință europeni, pe care aceștia au exprimat-o în mod clar la o întâlnire sub titlul semnificativ „Perspective pentru genomica în era post-genomică”, desfășurată la sfârșitul anului 2000 în Franța. Progresul științei americane în studierea genomului plantelor agricole și crearea formelor de plante transgenice, potrivit oamenilor de știință europeni, amenință că într-un viitor nu prea îndepărtat (două-cinci decenii), când creșterea populației va pune omenirea în fața unui criza alimentară, economia și știința europeană vor deveni dependente de tehnologia americană. În acest sens, a fost anunțată crearea unui program științific franco-german pentru studiul genomului plantelor („Plantgene”) și s-au făcut investiții semnificative în acesta.
Evident, problemele genomicei plantelor ar trebui să atragă atenția oamenilor de știință și organizatorilor ruși ai științei, precum și a organelor de conducere, deoarece nu este vorba doar de prestigiul științific, ci și de securitatea națională a țării. Într-un deceniu sau două, alimentele vor deveni cea mai importantă resursă strategică.
DIFICULTĂȚI ÎN STUDIAREA GENOMULUI PLANTELOR
Studiul genomului plantelor este o sarcină mult mai dificilă decât studiul genomului uman și al altor animale. Acest lucru se datorează următoarelor circumstanțe:
dimensiuni uriașe ale genomului, atingând zeci și chiar sute de miliarde de perechi de baze (pb) pentru speciile individuale de plante: genomul principalelor plante importante din punct de vedere economic (cu excepția orezului, inului și bumbacului) fie sunt apropiate ca mărime de genomul uman, fie depășește-l de multe ori (tabel);STUDII CROMOZOMIALE ALE GENOMURILORFluctuații puternice ale numărului de cromozomi la diferite plante - de la două la unele specii la câteva sute la altele și nu este posibil să se identifice o corelație strictă între dimensiunea genomului și numărul de cromozomi;
Se formează o abundență de poliploide (conținând mai mult de doi genomi per celulă) cu genomi similari, dar nu identici (alpoliploidie);
Îmbogățirea extremă a genomului plantelor (până la 99%) de ADN „nesemnificativ” (necodificare, adică neconținând gene), ceea ce face foarte dificilă unirea (aranjarea în ordinea corectă) a fragmentelor secvențiate într-un mod comun. regiune ADN de dimensiuni mari (contig);
Cartografierea morfologică, genetică și fizică incompletă a cromozomilor (comparativ cu genomul Drosophila, uman și șoarece);
Imposibilitatea practică a izolării cromozomilor individuali în formă pură folosind metode utilizate de obicei în acest scop pentru cromozomi umani și animale (sortarea în flux și utilizarea hibrizilor celulari);
Dificultatea cartografierii cromozomiale (determinarea locației pe cromozom) a genelor individuale folosind hibridizare in situ, datorită atât conținutului ridicat de ADN „nesemnificativ” din genomul plantelor, cât și particularităților organizării structurale a cromozomilor plantelor;
Depărtarea evolutivă a plantelor față de animale, ceea ce complică serios utilizarea informațiilor obținute prin secvențierea genomului oamenilor și a altor animale pentru studiul genomului plantelor;
Procesul lung de reproducere al majorității plantelor, care încetinește semnificativ analiza genetică a acestora.
Studiile cromozomiale (citogenetice) ale genomului în general și ale plantelor în special au o istorie lungă. Termenul „genom” a fost propus pentru a desemna un set haploid (unic) de cromozomi cu genele conținute în ei în primul sfert al secolului al XX-lea, adică cu mult înainte de stabilirea rolului ADN-ului ca purtător de informații genetice. .
Descrierea genomului unui organism multicelular nou, nestudiat genetic anterior, începe de obicei cu studiul și descrierea setului complet al cromozomilor săi (cariotip). Acest lucru, desigur, se aplică și plantelor, dintre care un număr mare nici măcar nu au început să fie studiate.
Deja în zorii studiilor cromozomiale, genomurile speciilor de plante înrudite au fost comparate pe baza analizei conjugării meiotice (combinație de cromozomi omologi) în hibrizi interspecifici. În ultimii 100 de ani, posibilitățile de analiză a cromozomilor s-au extins dramatic. Acum, pentru caracterizarea genomului plantelor sunt folosite tehnologii mai avansate: diverse variante ale așa-numitei colorări diferențiale, care face posibilă identificarea cromozomilor individuali după caracteristicile morfologice; hibridizare in situ făcând posibilă localizarea unor gene specifice pe cromozomi; studii biochimice ale proteinelor celulare (electroforeza si imunochimie) si, in final, un set de metode bazate pe analiza ADN-ului cromozomial pana la secventierea acestuia.

Orez. 1. Cariotipurile cerealelor a - secară (14 cromozomi), b - grâu dur (28 cromozomi), c - grâu moale (42 cromozomi), d - orz (14 cromozomi)De mulți ani, au fost studiate cariotipurile cerealelor, în primul rând grâul și secară. Interesant este că la diferite specii ale acestor plante, numărul de cromozomi este diferit, dar întotdeauna un multiplu de șapte. Tipurile individuale de cereale pot fi recunoscute în mod fiabil după cariotipul lor. De exemplu, genomul de secară este format din șapte perechi de cromozomi mari cu blocuri heterocromatice intens colorate la capete, adesea numite segmente sau benzi (Fig. 1a). Genomul grâului are deja 14 și 21 de perechi de cromozomi (Fig. 1, b, c), iar distribuția blocurilor heterocromatice în ele nu este aceeași ca și în cromozomii de secară. Genomii individuali de grâu, desemnați A, B și D, diferă, de asemenea, unul de celălalt. O creștere a numărului de cromozomi de la 14 la 21 duce la o schimbare bruscă a proprietăților grâului, care se reflectă în numele lor: durum, sau paste, grâu și moale, sau pâine, grâu . Gena D, care conține gene pentru proteinele glutenului, care conferă aluatului așa-numita germinație, este responsabilă pentru dobândirea de înalte proprietăți de coacere de către grâul moale. Acest genom este căruia i se acordă o atenție deosebită în îmbunătățirea selecției grâului de pâine. O altă cereală cu 14 cromozomi, orzul (Fig. 1, d), nu este de obicei folosită pentru a face pâine, dar este principala materie primă pentru fabricarea unor produse atât de comune precum berea și whisky-ul.
Cromozomii unor plante sălbatice utilizate pentru îmbunătățirea calității celor mai importante specii agricole, precum rudele sălbatice ale grâului - Aegilops, sunt în curs de studiu intens. Noi forme de plante sunt create prin încrucișare (Fig. 2) și selecție. În ultimii ani, o îmbunătățire semnificativă a metodelor de cercetare a făcut posibilă începerea studiului genomului plantelor, ale căror cariotipuri (în principal dimensiunea mică a cromozomilor) le făceau anterior inaccesibile pentru analiza cromozomilor. Așadar, abia recent au fost identificați pentru prima dată toți cromozomii de bumbac, mușețel și in.

Orez. 2. Cariotipuri de grâu și un hibrid de grâu cu Aegilops
a - grâu moale hexaploid ( Triticum astivum), constând din genomi A, B și O; b - grâu tetraploid ( Triticum timopheevi), constând din genomi A și G. conține gene de rezistență la majoritatea bolilor grâului; c - hibrizi Triticum astivum X Triticum timopheevi rezistent la mucegai și rugină, înlocuirea unei părți a cromozomilor este clar vizibilăSTRUCTURA PRIMARĂ A ADN-ului
Odată cu dezvoltarea geneticii moleculare, însuși conceptul de genom s-a extins. Acum acest termen este interpretat atât în sensul cromozomial clasic, cât și în sensul molecular modern: întregul material genetic al unui virus, al unei celule și al unui organism individual. Desigur, în urma studiului structurii primare complete a genomului (cum este adesea numită secvența liniară completă a bazelor de acid nucleic) a unui număr de microorganisme și oameni, a apărut problema secvențierii genomului plantelor.
Dintre numeroasele organisme vegetale, două au fost selectate pentru studiu - Arabidopsis, reprezentând clasa dicotiledoneelor (dimensiunea genomului 125 milioane bp) și orez din clasa monocotiledonelor (420-470 milioane bp). Acești genomi sunt mici în comparație cu alți genomi de plante și conțin relativ puține segmente ADN repetitive. Astfel de caracteristici au dat speranța că genomii selectați vor fi disponibili pentru determinarea relativ rapidă a structurii lor primare.

Orez. 3. Arabidopsis - muștar mic - o plantă mică din familia cruciferelor ( Brassicaceae). Pe un spațiu egal ca suprafață cu o pagină a revistei noastre, puteți crește până la o mie de organisme Arabidopsis individuale.Motivul alegerii Arabidopsis nu a fost doar dimensiunea redusă a genomului său, ci și dimensiunea redusă a organismului, ceea ce face ușoară creșterea lui în laborator (Fig. 3). Am luat în considerare ciclul său de reproducere scurt, datorită căruia este posibil să se efectueze rapid experimente privind încrucișarea și selecția, genetica studiată în detaliu, ușurința de manipulare în condițiile de creștere în schimbare (schimbarea compoziției de sare a solului, adăugarea diverșilor nutrienți etc. .) și testarea efectului asupra plantelor a diverșilor factori mutageni și agenți patogeni (viruși, bacterii, ciuperci). Arabidopsis nu are valoare economică, prin urmare, genomul său, împreună cu genomul șoarecelui, a fost numit referință sau, mai puțin precis, model.*
* Apariția termenului „genom model” în literatura rusă este rezultatul unei traduceri inexacte a expresiei engleze model genom. Cuvântul „model” înseamnă nu numai adjectivul „model”, ci și substantivul „probă”, „standard”, „model”. Ar fi mai corect să vorbim despre un genom eșantion sau despre un genom de referință.Lucrările intensive privind secvențierea genomului Arabidopsis au fost începute în 1996 de către un consorțiu internațional care includea instituții științifice și grupuri de cercetare din SUA, Japonia, Belgia, Italia, Marea Britanie și Germania. În decembrie 2000, au devenit disponibile informații extinse care rezumă determinarea structurii primare a genomului Arabidopsis. Pentru secvențiere a fost folosită tehnologia clasică sau ierarhică: mai întâi, au fost studiate secțiuni individuale mici ale genomului, din care au fost compuse secțiuni mai mari (contigs), iar în etapa finală, structura cromozomilor individuali. ADN-ul nuclear al genomului Arabidopsis este distribuit pe cinci cromozomi. În 1999, au fost publicate rezultatele secvențierii a doi cromozomi, iar apariția în presă a informațiilor despre structura primară a celor trei rămași a completat secvențierea întregului genom.
Din 125 de milioane de perechi de baze, a fost determinată structura primară de 119 milioane, ceea ce reprezintă 92% din întregul genom. Doar 8% din genomul Arabidopsis care conține blocuri mari de segmente repetitive de ADN s-a dovedit a fi inaccesibil pentru studiu. În ceea ce privește completitatea și minuțiozitatea secvențierii genomului eucariotic, Arabidopsis rămâne încă în primii trei campioni împreună cu un organism de drojdie unicelulară. Saccharomyces cerevisiaeși organism multicelular Caenorhabditis eleganta(Vezi tabelul).
Aproximativ 15.000 de gene individuale care codifică proteine au fost găsite în genomul Arabidopsis. Aproximativ 12 000 dintre acestea sunt conținute sub formă de două copii per genom haploid (unic), astfel încât numărul total de gene este de 27 000. Numărul de gene din Arabidopsis nu diferă mult de numărul de gene din organisme precum oamenii și șoarecii, dar dimensiunea genomului său de 25-30 de ori mai mică. Această împrejurare este asociată cu caracteristici importante în structura genelor individuale Arabidopsis și structura generală a genomului său.
Genele Arabidopsis sunt compacte, conținând doar câțiva exoni (regiuni care codifică proteine) separați de segmente scurte (aproximativ 250 bp) de ADN necodificatori (introni). Intervalele dintre genele individuale sunt în medie de 4600 de perechi de baze. Pentru comparație, subliniem că genele umane conțin multe zeci și chiar sute de exoni și introni, iar regiunile intergenice au dimensiuni de 10 mii de perechi de baze sau mai mult. Se presupune că prezența unui genom compact mic a contribuit la stabilitatea evolutivă a Arabidopsis, deoarece ADN-ul său a devenit o țintă pentru diferiți agenți dăunători într-o măsură mai mică, în special pentru introducerea de fragmente de ADN repetitive asemănătoare virusului (transpozoni). în genom.
Printre alte caracteristici moleculare ale genomului Arabidopsis, trebuie remarcat faptul că exonii sunt îmbogățiți în guanină și citozină (44% în exoni și 32% în introni) în comparație cu genele animale, precum și prezența genelor dublu repetate (duplicate). Se presupune că o astfel de dublare a avut loc ca urmare a a patru evenimente simultane, constând în dublarea (repetarea) unei părți a genelor Arabidopsis sau fuziunea genomilor înrudite. Aceste evenimente, care au avut loc în urmă cu 100-200 de milioane de ani, sunt o manifestare a tendinței generale de poliploidizare (o creștere multiplă a numărului de genomi dintr-un organism), care este caracteristică genomului plantelor. Cu toate acestea, unele fapte arată că genele duplicate în Arabidopsis nu sunt identice și funcționează diferit, ceea ce poate fi asociat cu mutații în regiunile lor de reglementare.
Orezul a devenit un alt obiect de secvențiere completă a ADN-ului. Genomul acestei plante este, de asemenea, mic (12 cromozomi, dând un total de 420-470 milioane bp), doar de 3,5 ori mai mare decât cel al Arabidopsis. Cu toate acestea, spre deosebire de Arabidopsis, orezul are o importanță economică deosebită, fiind baza alimentației pentru mai mult de jumătate din umanitate, prin urmare, nu numai miliarde de consumatori, ci și o armată de oameni multimilionare implicați activ în procesul foarte laborios al acestuia. cultivarea sunt extrem de interesați să-și îmbunătățească proprietățile.
Unii cercetători au început să studieze genomul orezului încă din anii 1980, dar aceste studii au atins o amploare serioasă abia în anii 1990. În 1991, în Japonia a fost creat un program de descifrare a structurii genomului orezului, reunind eforturile multor grupuri de cercetare. În 1997, pe baza acestui program a fost organizat Proiectul Internațional al Genomului Orezului. Participanții săi au decis să-și concentreze eforturile pe secvențierea uneia dintre subspeciile de orez ( Oriza sativajaponica), în studiul căruia s-au înregistrat deja progrese semnificative până la acel moment. Un stimulent serios și, la figurat vorbind, o vedetă călăuzitoare pentru o astfel de muncă a fost programul „Genom uman”.
În cadrul acestui program a fost testată strategia de diviziune ierarhică „cromozomială” a genomului, pe care participanții consorțiului internațional au folosit-o pentru a descifra genomul orezului. Cu toate acestea, dacă în studiul genomului uman, fracțiile de cromozomi individuali au fost izolate folosind diverse metode, atunci materialul specific pentru cromozomii individuali de orez și regiunile lor individuale a fost obținut prin microdisecție cu laser (decuparea obiectelor microscopice). Pe o lamă de microscop, unde se află cromozomii de orez, sub influența unui fascicul laser, totul este ars, cu excepția cromozomului sau a secțiunilor acestuia programate pentru analiză. Materialul rămas este folosit pentru clonare și secvențiere.
Au fost publicate numeroase rapoarte cu privire la rezultatele secvențierii fragmentelor individuale ale genomului orezului, efectuate cu mare acuratețe și detaliu, caracteristice tehnologiei ierarhice. Se credea că determinarea structurii primare complete a genomului orezului va fi finalizată până la sfârșitul anului 2003 – mijlocul anului 2004, iar rezultatele, împreună cu datele privind structura primară a genomului Arabidopsis, vor fi utilizate pe scară largă în comparație. genomica altor plante.
Cu toate acestea, la începutul anului 2002, două grupuri de cercetare - unul din China, celălalt din Elveția și Statele Unite - au publicat rezultatele unui proiect complet de secvențiere (aproximativă) a genomului orezului, realizat folosind tehnologia de clonare totală. Spre deosebire de studiul în etape (ierarhice), abordarea totală se bazează pe clonarea simultană a întregului ADN genomic într-unul dintre vectorii virali sau bacterieni și obținerea unui număr semnificativ (uriaș pentru genomi medii și mari) de clone individuale care conțin diverse Segmente de ADN. Pe baza analizei acestor secțiuni secvențiate și a suprapunerii secțiunilor terminale identice ale ADN-ului, se formează un contig - un lanț de secvențe ADN unite între ele. Contigul general (total) este structura primară a întregului genom sau cel puțin a unui cromozom individual.
Într-o astfel de prezentare schematică, strategia clonării totale pare simplă. De fapt, întâmpină dificultăți serioase asociate cu necesitatea obținerii unui număr imens de clone (se acceptă în general că genomul sau regiunea sa studiată trebuie acoperite de clone de cel puțin 10 ori), o cantitate imensă de secvențiere și extrem de complexă. munca la clone de andocare care necesită participarea specialiștilor în bioinformatică. Un obstacol serios în calea clonării totale este o varietate de segmente ADN repetitive, al căror număr, după cum sa menționat deja, crește brusc pe măsură ce dimensiunea genomului crește. Prin urmare, strategia de secvențiere totală este utilizată în principal în studiul genomului virusurilor și microorganismelor, deși a fost folosită cu succes pentru a studia genomul unui organism multicelular, Drosophila.
Rezultatele secvențierii totale a acestui genom au fost „suprapuse” unei game uriașe de informații despre structura sa cromozomială, genică și moleculară, obținute pe o perioadă de aproape 100 de ani de studiu a Drosophila. Și totuși, în ceea ce privește gradul de secvențiere, genomul Drosophila (66% din dimensiunea totală a genomului) este semnificativ inferior genomul Arabidopsis (92%), în ciuda dimensiunilor lor destul de apropiate - 180 de milioane și, respectiv, 125 de milioane de perechi de baze. . Prin urmare, recent s-a propus denumirea tehnologiei mixte, care a fost folosită pentru secvențierea genomului Drosophila.
Pentru a secvenționa genomul orezului, grupurile de cercetare menționate mai sus au luat două dintre subspeciile sale, cele mai cultivate în țările asiatice, - Oriza saliva L. ssp indicajȘi Oriza saliva L. sspjaponica. Rezultatele studiilor lor coincid în multe privințe, dar diferă în multe privințe. Astfel, reprezentanții ambelor grupuri au declarat că au ajuns la aproximativ 92-93% din suprapunerea genomului cu contigs. S-a demonstrat că aproximativ 42% din genomul orezului este reprezentat de repetări scurte de ADN constând din 20 de perechi de baze, iar majoritatea elementelor mobile de ADN (transpozoni) sunt localizate în regiuni intergenice. Cu toate acestea, datele privind dimensiunea genomului orezului diferă semnificativ.
Pentru subspecia japoneză, dimensiunea genomului este determinată a fi de 466 milioane de perechi de baze, iar pentru subspecia indiană, 420 milioane. Motivul acestei discrepanțe nu este clar. Poate fi o consecință a diferitelor abordări metodologice în determinarea dimensiunii părții necodificatoare a genomului, adică nu reflectă adevărata stare de lucruri. Dar este posibil să existe o diferență de 15% în dimensiunea genomilor studiati.
A doua discrepanță majoră a fost dezvăluită în numărul de gene găsite: pentru subspecia japoneză, de la 46 022 la 55 615 gene per genom, iar pentru subspecia indiană, de la 32 000 la 50 000. Motivul acestei discrepanțe nu este clar.
Incompletitudinea și inconsecvența informațiilor primite se remarcă în comentariile la articolele publicate. De asemenea, aici se exprimă speranța că lacunele în cunoașterea genomului orezului vor fi eliminate prin compararea datelor de „secvențiere brută” cu rezultatele secvențierii detaliate, ierarhice, efectuate de participanții la Proiectul Internațional de Genom al Orezului.
GENOMIA PLANTELOR COMPARATIVA SI FUNCTIONALA
Datele extinse obținute, dintre care jumătate (rezultatele grupului chinez) sunt disponibile publicului, deschid fără îndoială perspective largi atât pentru studiul genomului orezului, cât și pentru genomica plantelor în general. O comparație a proprietăților genomului Arabidopsis și al orezului a arătat că majoritatea genelor (până la 80%) identificate în genomul Arabidopsis se găsesc și în genomul orezului, cu toate acestea, pentru aproximativ jumătate din genele găsite în orez, analogi ( ortologi) nu au fost încă găsite în genomul Arabidopsis. În același timp, 98% dintre genele a căror structură primară a fost stabilită pentru alte cereale au fost găsite în genomul orezului.
Discrepanța semnificativă (aproape dublă) dintre numărul de gene din orez și Arabidopsis este uluitoare. În același timp, datele proiectului de decodificare a genomului orezului, obținute prin secvențierea totală, practic nu sunt comparate cu rezultatele ample ale studiului genomului orezului prin metoda clonării și secvențierii ierarhice, adică ceea ce are nu a fost efectuată cu privire la genomul Drosophila. Prin urmare, rămâne neclar dacă diferența în numărul de gene în Arabidopsis și orez reflectă adevărata stare de lucruri sau dacă este explicată prin diferența de abordări metodologice.
Spre deosebire de genomul Arabidopsis, nu sunt date date despre genele gemene din genomul orezului. Este posibil ca cantitatea lor relativă să fie mai mare în orez decât în Arabidopsis. Această posibilitate este susținută indirect de date privind prezența formelor poliploide de orez. Se poate aștepta mai multă claritate cu privire la această problemă după finalizarea Proiectului Internațional de Genom al Orezului și se obține o imagine detaliată a structurii primare a ADN-ului acestui genom. Motive serioase pentru o astfel de speranță sunt oferite de faptul că, după publicarea lucrărilor privind secvențierea brută a genomului orezului, numărul de publicații privind structura acestui genom a crescut brusc, în special, au apărut informații despre secvențierea detaliată. dintre cromozomii 1 și 4 ai săi.
Cunoașterea, cel puțin aproximativ, a numărului de gene din plante este de o importanță fundamentală pentru genomica comparativă a plantelor. Inițial, s-a crezut că, deoarece toate plantele cu flori sunt foarte apropiate una de cealaltă în ceea ce privește caracteristicile lor fenotipice, genomul lor ar trebui să fie, de asemenea, similar. Și dacă studiem genomul Arabidopsis, vom obține informații despre majoritatea genomilor altor plante. O confirmare indirectă a acestei presupuneri sunt rezultatele secvențierii genomului șoarecelui, care este surprinzător de aproape de genomul uman (aproximativ 30 de mii de gene, dintre care doar 1 mie s-au dovedit a fi diferite).
Se poate presupune că motivul diferențelor dintre genomul Arabidopsis și orezul constă în apartenența lor la diferite clase de plante - dicotiledone și monocotiledone. Pentru a clarifica această problemă, este foarte de dorit să se cunoască cel puțin o structură primară brută a unei alte plante monocotiledonate. Cel mai realist candidat ar putea fi porumbul, al cărui genom este aproximativ egal cu genomul uman, dar totuși mult mai mic decât genomul altor cereale. Valoarea nutritivă a porumbului este binecunoscută.
Vastul material obținut ca urmare a secvențialării genomului Arabidopsis și orezului devine treptat baza unui studiu la scară largă al genomului plantelor folosind genomica comparativă. Astfel de studii au o semnificație biologică generală, deoarece fac posibilă stabilirea principiilor principale ale organizării genomului plantei în ansamblu și a cromozomilor lor individuali, identificarea trăsăturilor comune ale structurii genelor și regiunilor lor de reglare și luarea în considerare a acestora. raportul dintre partea activă funcțional (genă) a cromozomului și diferitele regiuni ADN intergenice care nu codifică proteine. Genetica comparativă devine, de asemenea, din ce în ce mai importantă pentru dezvoltarea genomicii funcționale umane. Pentru studii comparative s-a efectuat secvențierea genomilor de pește-papă și șoarece.
La fel de important este studiul genelor individuale responsabile de sinteza proteinelor individuale care determină funcțiile specifice ale corpului. Este în descoperirea, izolarea, secvențierea și determinarea funcției genelor individuale în care se află semnificația practică, în primul rând medicală, a programului Genomului uman. Această împrejurare a fost remarcată în urmă cu câțiva ani de către J. Watson, care a subliniat că programul Genomului uman va fi finalizat numai atunci când vor fi determinate funcțiile tuturor genelor umane.

Orez. 4. Clasificare în funcție de funcția genelor Arabidopsis
1 - gene pentru creștere, diviziune și sinteza ADN; 2 - gene de sinteză a ARN (transcripție); 3 - gene pentru sinteza si modificarea proteinelor; 4 - gene pentru dezvoltare, îmbătrânire și moarte celulară; 5 - gene ale metabolismului celular și ale metabolismului energetic; 6 - gene de interacțiune intercelulară și transmitere a semnalului; 7 - gene pentru furnizarea altor procese celulare; 8 - gene cu funcție necunoscutăÎn ceea ce privește funcția genelor plantelor, știm mai puțin de o zecime din ceea ce știm despre genele umane. Chiar și în Arabidopsis, al cărui genom este mult mai studiat decât genomul uman, funcția a aproape jumătate din genele sale rămâne necunoscută (Fig. 4). Între timp, pe lângă genele comune animalelor, plantele au un număr semnificativ de gene care sunt specifice doar (sau cel puțin predominant) pentru acestea. Vorbim despre genele implicate în transportul apei și sinteza peretelui celular, care este absent la animale, despre genele care asigură formarea și funcționarea cloroplastelor, fotosinteza, fixarea azotului, precum și sinteza a numeroase produse aromatice. Această listă poate fi continuată, dar este deja clar cu ce sarcină dificilă se confruntă genomica funcțională a plantelor.
Secvențierea întregului genom oferă informații aproape adevărate despre numărul total de gene dintr-un organism dat, face posibilă plasarea unor informații mai mult sau mai puțin detaliate și fiabile despre structura lor în băncile de date și facilitează munca de izolare și studiere a genelor individuale. Cu toate acestea, secvențierea genomului nu înseamnă în niciun caz să stabilească funcția tuturor genelor.
Una dintre cele mai promițătoare abordări ale genomicii funcționale se bazează pe identificarea genelor de lucru care sunt utilizate pentru transcrierea (citirea) ARNm. Această abordare, inclusiv utilizarea tehnologiei moderne de microarray, face posibilă identificarea simultană a până la zeci de mii de gene funcționale. Recent, folosind această abordare, a început studiul genomului plantelor. Pentru Arabidopsis, a fost posibil să se obțină aproximativ 26 de mii de transcrieri individuale, ceea ce facilitează foarte mult posibilitatea de a determina funcția aproape tuturor genelor sale. La cartofi, a fost posibil să se identifice aproximativ 20.000 de gene de lucru care sunt importante pentru înțelegerea atât a proceselor de creștere și formare a tuberculilor, cât și a proceselor de boală a cartofului. Se presupune că aceste cunoștințe vor crește rezistența unuia dintre cele mai importante produse alimentare la agenți patogeni.
Dezvoltarea logică a genomicii funcționale a fost proteomica. Acest nou domeniu de știință studiază proteomul, care este de obicei înțeles ca un set complet de proteine dintr-o celulă la un anumit moment. Un astfel de set de proteine, care reflectă starea funcțională a genomului, se schimbă tot timpul, în timp ce genomul rămâne neschimbat.
Studiul proteinelor a fost folosit de mult timp pentru a judeca activitatea genomului plantelor. După cum se știe, enzimele prezente în toate plantele diferă în fiecare specie și soiuri în secvența de aminoacizi. Astfel de enzime, cu aceeași funcție, dar cu o secvență diferită de aminoacizi individuali, se numesc izoenzime. Au proprietăți fizico-chimice și imunologice diferite (greutate moleculară, sarcină), care pot fi detectate prin cromatografie sau electroforeză. De mulți ani, aceste metode au fost folosite cu succes pentru a studia așa-numitul polimorfism genetic, adică diferențele dintre organisme, soiuri, populații, specii, în special grâul și formele înrudite de cereale. Recent, însă, datorită dezvoltării rapide a metodelor de analiză a ADN-ului, inclusiv secvențierea, studiul polimorfismului proteic a fost înlocuit cu studiul polimorfismului ADN. Cu toate acestea, studiul direct al spectrelor proteinelor de depozitare (prolamine, gliadine etc.), care determină principalele proprietăți nutriționale ale cerealelor, rămâne o metodă importantă și de încredere pentru analiza genetică, selecția și producția de semințe a plantelor agricole.
Cunoașterea genelor, a mecanismelor de exprimare și reglare a acestora este extrem de importantă pentru dezvoltarea biotehnologiei și producerea de plante transgenice. Se știe că succesele impresionante în acest domeniu provoacă o reacție ambiguă din partea comunității de mediu și medicale. Cu toate acestea, există o zonă a biotehnologiei vegetale în care aceste temeri, dacă nu complet nefondate, atunci, în orice caz, par a fi de puțină importanță. Vorbim despre crearea de plante industriale transgenice care nu sunt folosite ca produse alimentare. India a recoltat recent prima recoltă de bumbac transgenic care este rezistent la o serie de boli. Există informații despre introducerea unor gene speciale care codifică proteine pigmentare în genomul bumbacului și producerea de fibre de bumbac care nu necesită vopsire artificială. O altă cultură industrială care poate face obiectul unei inginerie genetică eficientă este inul. Utilizarea sa ca alternativă la bumbac pentru materii prime textile a fost discutată recent. Această problemă este extrem de importantă pentru țara noastră, care și-a pierdut propriile surse de bumbac brut.
PERSPECTIVE DE STUDIAREA GENOMULUI PLANTELOR
Evident, studiile structurale ale genomului plantelor se vor baza pe abordările și metodele genomicei comparative, folosind ca material principal rezultatele descifrării genomului Arabidopsis și orezului. Un rol important în dezvoltarea genomicii comparative a plantelor îl vor juca, fără îndoială, informațiile care mai devreme sau mai târziu vor fi furnizate de secvențierea totală (aproximată) a genomilor altor plante. În acest caz, genomica comparativă a plantelor se va baza pe stabilirea de relații genetice între loci individuali și cromozomi aparținând unor genomi diferiți. Ne vom concentra nu atât pe genomica generală a plantelor, cât pe genomica selectivă a locurilor cromozomiale individuale. De exemplu, s-a demonstrat recent că gena responsabilă pentru vernalizare este localizată la locusul VRn-AI al cromozomului hexaploid de grâu 5A și locusul Hd-6 al cromozomului 3 de orez.
Dezvoltarea acestor studii va fi un impuls puternic pentru identificarea, izolarea și secvențierea multor gene ale plantelor importante din punct de vedere funcțional, în special, genele responsabile de rezistența la boli, rezistența la secetă și adaptabilitatea la diferite condiții de creștere. Din ce în ce mai mult, se va folosi genomica funcțională, bazată pe detectarea în masă (screening) a genelor care funcționează în plante.
Putem prevedea îmbunătățiri suplimentare ale tehnologiilor cromozomiale, în primul rând metoda de microdisecție. Utilizarea sa extinde dramatic posibilitățile cercetării genomice fără a necesita costuri uriașe, cum ar fi, de exemplu, secvențierea totală a genomului. Metoda de localizare pe cromozomii plantelor a genelor individuale cu ajutorul hibridizării va fi răspândită în continuare. in situ.În prezent, utilizarea sa este limitată de numărul mare de secvențe repetitive din genomul plantei și, eventual, de particularitățile organizării structurale a cromozomilor plantelor.
Tehnologiile cromozomiale vor deveni de mare importanță pentru genomica evolutivă a plantelor în viitorul apropiat. Aceste tehnologii relativ ieftine fac posibilă evaluarea rapidă a variabilității intra și interspecifice, studiul genomilor alopoliploizi complexe de grâu tetraploid și hexaploid, triticale; analiza proceselor evolutive la nivel cromozomial; investighează formarea genomilor sintetici și introducerea (introgresiunea) materialului genetic străin; identificarea relațiilor genetice dintre cromozomii individuali ai diferitelor specii.
Studiul cariotipului plantelor folosind metode citogenetice clasice, îmbogățit prin analiză biologică moleculară și tehnologie computerizată, va fi utilizat pentru caracterizarea genomului. Acest lucru este deosebit de important pentru studierea stabilității și variabilității cariotipului la nivelul nu numai al organismelor individuale, ci și al populațiilor, soiurilor și speciilor. În cele din urmă, este dificil de imaginat cum poate fi estimat numărul și spectrele de rearanjamente cromozomiale (aberații, punți) fără utilizarea metodelor de colorare diferențială. Astfel de studii sunt extrem de promițătoare pentru monitorizarea mediului în funcție de starea genomului plantei.
În Rusia modernă, secvențierea directă a genomului plantelor este puțin probabil să fie efectuată. O astfel de muncă, care necesită investiții mari, depășește puterea economiei noastre actuale. Între timp, datele privind structura genomului Arabidopsis și orezului, obținute de știința mondială și disponibile în băncile internaționale de date, sunt suficiente pentru dezvoltarea genomicii plantelor interne. Putem prevedea extinderea studiilor genomului plantelor bazate pe abordări de genomică comparativă pentru a rezolva probleme specifice de ameliorare și producție a culturilor, precum și pentru a studia originea diferitelor specii de plante de mare importanță economică.
Se poate presupune că abordările genomice precum tiparea genetică (analize RELF, RAPD, AFLP etc.), care sunt destul de accesibile pentru bugetul nostru, vor fi utilizate pe scară largă în practica internă de ameliorare și producția de culturi. În paralel cu metodele directe de determinare a polimorfismului ADN, abordări bazate pe studiul polimorfismului proteic, în primul rând proteinele de depozitare a cerealelor, vor fi utilizate pentru rezolvarea problemelor de genetică și ameliorare a plantelor. Tehnologiile cromozomiale vor fi utilizate pe scară largă. Sunt relativ ieftine, dezvoltarea lor necesită investiții destul de moderate. În domeniul studiilor cromozomiale, știința domestică nu este inferioară lumii.
Trebuie subliniat faptul că știința noastră a adus o contribuție semnificativă la formarea și dezvoltarea genomicii plantelor [ , ].
Rolul fundamental l-a jucat N.I. Vavilov (1887-1943).
În biologia moleculară și genomica plantelor, contribuția de pionierat a lui A.N. Belozerski (1905-1972).
În domeniul studiilor cromozomiale, este de remarcat munca remarcabilului genetician S.G. Navashin (1857-1930), care a descoperit primul cromozomi sateliti la plante și a demonstrat că este posibil să se facă distincția între cromozomi individuali în funcție de caracteristicile morfologiei lor.
Un alt clasic al științei ruse G.A. Levitsky (1878-1942) a descris în detaliu cromozomii de secară, grâu, orz, mazăre și sfeclă de zahăr, a introdus termenul „cariotip” în știință și a dezvoltat doctrina acestuia.
Specialiștii moderni, bazându-se pe realizările științei mondiale, pot aduce o contribuție semnificativă la dezvoltarea ulterioară a geneticii și genomicii plantelor.
Autorul își exprimă sincere mulțumiri academicianului Yu.P. Altukhov pentru discuția critică a articolului și sfaturi valoroase.Activitatea echipei conduse de autorul articolului a fost susținută de Fundația Rusă pentru Cercetare de bază (granturi nr. 99-04-48832; 00-04-49036; 00-04-81086), Programul președintelui Federația Rusă pentru a sprijini școlile științifice (granturile nr. 00-115 -97833 și NSh-1794.2003.4) și Programul Academiei Ruse de Științe „Markeri genetici moleculari și cromozomiali în dezvoltarea metodelor moderne de reproducere și producție de semințe” .
LITERATURĂ
1. Zelenin A.V., Badaeva E.D., Muravenko O.V. Introducere în genomica plantelor // Biologie moleculară. 2001. V. 35. S. 339-348. 2. Pen E. Bonanza pentru genomica plantelor // Știință. 1998. V. 282. P. 652-654. 3. Genomica plantelor, Proc. Natl. Acad. sci. STATELE UNITE ALE AMERICII. 1998. V. 95. P. 1962-2032. 4. Cartel N.A. si etc. Genetica. Dicţionar enciclopedic. Minsk: Technologia, 1999. 5. Badaeva E.D., Friebe B., Gill B.S. 1996. Diferențierea genomului în Aegilops. 1. Distribuția secvențelor de ADN foarte repetitive pe cromozomii speciilor diploide // Genom. 1996. V. 39. P. 293-306. Istoricul analizei cromozomilor // Biol. membranelor. 2001. T. 18. S. 164-172.




