Кои водорасли нямат флагеларни стадии. Нисши растения или водорасли. Самотни свободно живеещи форми
Монадичните вегетативни клетки и монадичните етапи в жизнения цикъл (зооспори и гамети) на водораслите са снабдени с флагели - дълги и доста дебели израстъци на клетки, външно покрити с плазмалема. Техният брой, дължина, морфология, място на прикрепване, характер на движение са доста разнообразни при водораслите, но са постоянни в рамките на сродни групи.
Камшичетата могат да бъдат прикрепени към предния край на клетката (апикално) или могат да бъдат леко изместени настрани (субапикално); тяхното закрепване е възможно отстрани на клетката (латерално) и от вентралната страна на клетката (вентрално). Наричат се флагели, които са идентични по морфология изоморфенако се различават - хетероморфен. Изоконт- това са флагели с еднаква дължина, хетероконтакт- различни дължини.
Камшичетата имат единен структурен план. Възможно е да се разграничи свободната част (ундулиподия), преходната зона, базалното тяло (кинетозома). Различните части на флагела се различават по броя и разположението на микротубулите, които образуват скелета (фиг. 2).
Ориз. Фиг. 2. Схема на структурата на флагела на водорасли (според: L.L. Velikanov et al., 1981): 1 - надлъжен разрез на флагела; 2, 3 - напречен разрез през върха на флагела; 4 - напречно сечение през undulipodia; 5 - преходна зона; 6 - напречно сечение през основата на флагела - кинетозома
Ундулиподий(в превод от латински "waveleg") е способен да извършва ритмични вълнообразни движения. Undulipodium е покрита с мембрана аксонема. аксонемасе състои от девет двойки микротубули, подредени в кръг и двойка микротубули в центъра (фиг. 2). Камшичетата могат да бъдат гладки или покрити с люспи или мастигонеми (косми), докато при динофитите и криптофитите те са покрити както с люспи, така и с косми. Камшичетата на примизиофитите, криптофитите и зелените водорасли могат да бъдат покрити с люспи с различни форми и размери.
преходна зона. Функционално играе роля в укрепването на флагела на мястото на излизането му от клетката. При водораслите се разграничават няколко вида структури на преходната зона: напречна плоча (динофити), звездовидна структура (зелена), преходна спирала (хетероконт), преходен цилиндър (примнезиофити и динофити).
Базално тяло или кинетозома. Тази част от флагела има структура под формата на кух цилиндър, чиято стена е образувана от девет триплета микротубули. Функцията на кинетозомата е връзката на флагела с плазмалемата на клетката. Базалните тела в редица водорасли могат да участват в ядреното делене и да станат центрове на микротубулна организация.
Край на работата -
Тази тема принадлежи на:
Морски водорасли
Рибарски университет.. Институт по морска биология, кръстен на v Zhirmunsky Dvor.
Ако имате нужда от допълнителен материал по тази тема или не сте намерили това, което търсите, препоръчваме да използвате търсенето в нашата база данни с произведения:
Какво ще правим с получения материал:
Ако този материал се оказа полезен за вас, можете да го запазите на страницата си в социалните мрежи:
| туит |
Всички теми в този раздел:
Калъфи за клетки
Клетъчните покрития осигуряват устойчивост на вътрешното съдържание на клетките на външни влияния и придават на клетките определена форма. Обвивките са пропускливи за вода и ниски молекули, разтворени в нея
Митохондриите
Митохондриите се намират в клетките на еукариотните водорасли. Формата и структурата на митохондриите в клетките на водораслите са по-разнообразни, отколкото в митохондриите на висшите растения. Може да са кръгли.
Пигменти
Всички водорасли се отличават добре с набор от фотосинтетични пигменти. Такива групи в таксономията на растенията имат статут на отдели. Основният пигмент на всички водорасли е зеленият пигмент.
пластиди
Пигментите в клетките на еукариотните водорасли се намират в пластидите, както при всички растения. Има два вида пластиди във водораслите: цветни хлоропласти (хроматофори) и безцветни левкопласти (ami
Ядро и митотичен апарат
Ядрото на водораслите има структура, характерна за еукариотите. Броят на ядрата в една клетка може да варира от едно до няколко. Отвън ядрото е покрито с мембрана, състояща се от две мембрани, външната мембрана
Монадичен (флагелиран) тип структура на талуса
Най-характерната особеност, която определя този тип структура, е наличието на флагели, с помощта на които монадните организми се движат активно във водната среда (фиг. 9, А). Подвижна ж
Ризоподиален (амебоиден) тип структура
Най-важните характеристики на амебоидния тип структура са липсата на здрави клетъчни обвивки и способността за амебоидно движение, с помощта на временно образуваната на повърхността на клетката ци.
Палмелоиден (хемимонас) тип структура
Характерно за този тип структура е комбинацията от неподвижен начин на живот на растенията с наличието на клетъчни органели, характерни за монадичните организми: контрактилни вакуоли, стигма, турникет
Кокоиден тип структура
Този тип съчетава едноклетъчни и колониални водорасли, неподвижни във вегетативно състояние. Клетките от кокоиден тип са покрити с мембрана и имат протопласт от растителен тип (тонопласт без сократ).
Трихален (нишковиден) тип структура
Характерна особеност на нишковидния тип структура е нишковидното разположение на неподвижни клетки, които се образуват вегетативно в резултат на клетъчното делене, което се случва предимно
Хетеротрихален (многофилен) тип структура
Мултифиламентният тип възниква на базата на нишковидния тип. Многовлакнестият талус се състои предимно от хоризонтални нишки, които се простират по протежение на субстрата, които изпълняват функцията на прикрепване, и вертикални, по
Паренхимен (тъканен) тип структура
Една от посоките на еволюцията на мултифиламентния талус е свързана с появата на паренхимни тали. Способността за неограничен растеж и делене на клетките в различни посоки доведе до образуването
Структура тип сифон
Сифонният (неклетъчен) тип структура се характеризира с липсата на талус вътре, който достига относително големи, обикновено макроскопични размери и определена степен на диференциация, клетки
Тип конструкция със сифон
Основната характеристика на сифонния тип структура е способността да се образуват сложни тали от първичния неклетъчен талус, състоящ се от първични многоядрени сегменти. IN
безполово размножаване
Безполовото размножаване на водораслите се осъществява с помощта на специализирани клетки - спори. Спорулацията обикновено се придружава от разделянето на протопласта на части и освобождаването на продукти на делене от
просто разделение
Този метод на размножаване се среща само при едноклетъчни форми на водорасли. Най-простото делене се случва в клетки, които имат амебоиден тип структура на тялото. Разделяне на амебоидните форми
Раздробяване
Фрагментацията е присъща на всички групи многоклетъчни водорасли и се проявява в различни форми: образуване на хормони, регенерация на откъснати части от талуса, спонтанно падане на клони, повторно израстване
Размножаване чрез издънки, столони, плодови пъпки, възли, акинети
В тъканните форми на зелени, кафяви и червени водорасли вегетативното размножаване придобива пълна форма, която малко се различава от вегетативното размножаване на висшите растения. Поддържане на пътя
полово размножаване
полово размножаванепри водораслите се свързва със сексуалния процес, който се състои в сливане на две клетки, което води до образуването на зигота, която расте в нов индивид или дава зооспори.
Промяна на ядрените фази
По време на половия процес, в резултат на сливането на гамети и техните ядра, броят на хромозомите в ядрото се удвоява. На определен етап от цикъла на развитие, по време на мейозата, настъпва намаляване на броя на хромозомите, в резултат на
Ендофити / ендозоити или ендосимбионти
Ендосимбионтите или вътреклетъчните симбионти са водорасли, които живеят в тъканите или клетките на други организми (безгръбначни или водорасли). Те образуват своеобразна екологична група.
Отдел синьо-зелени водорасли (цианобактерии) - cyanophyta
Името на отдела (от гръцки cyanos - синьо) отразява характерна особеност на тези водорасли - цвета на талуса, свързан с относително високото съдържание на синия пигмент фикоцианин. Цианоген
Разред - Chroococcales
Те се срещат като едноклетъчни "прости" индивиди или по-често образуват лигавични колонии. Когато клетките се разделят в две равнини, се появяват еднослойни ламеларни колонии. Разделяне на три n
Отдел Червени водорасли - родофити
Името на катедрата идва от гръцка думародон ("радон") - розов. Цветът на червените водорасли се дължи на различна комбинация от пигменти. Тя идва от сиво и лилаво
Разред Bangiaceae–Bangiales
Родът Porphyra има талус под формата на тънка лъскава плоча с гладки или нагънати ръбове, състояща се от един или два слоя плътно свързани клетки. Основата на чинията обикновено влиза в
Разред Rhodimeniales - Rhodymeniales
Род Sparlingia (Rhodimenia) - плоски плочи до 45 см височина, листовидни и клиновидни, разширени и дланово разчленени на върха, от светло розово или светло оранжево до
Разред Коралови - Corallinales
Род Corallina - съединени ветрилообразни разклонени храсти до 10 см височина, разклонени, варовити, от розово-люляк до почти бяло. Размножава се безполово и полово. Спо
Гигартински ред - Gigartinales
Rod Chondrus - плътни кожести хрущялни храсти до 20 см височина, 3-4 пъти разклонени, светложълти, светло розови, лилаво-тъмночервени. Расте в долната част на крайбрежието и
Разред Керамиеви - Ceramiales
Род Ceramium - нежни пухкави съчленени храсти до 10 см височина, дихотомно или последователно разклонени, тъмно жълти с розов оттенък. Разклонение от два до четири реда, крайно
Отдел диатомеи - bacilariophyta
Отделът се нарича Диатомеи (от гръцки di - две, tome - рязане, дисекция) или Bacillaria (bacillum - пръчка). Включва едноклетъчни самотни или колониални орг
Отдел хетероконтни (многофлагелатни) водорасли - heterokontophyta
Всички хетероконти изглежда имат флагеларен апарат. Камшичетата са 2, като на единия от тях има много характерни тръбести тричленни перести израстъци, или власинки - мастигонеми. Това е пари в брой
Систематика
Фосилните коколити са известни от мезозойските отлагания и са били в изобилие през по-голямата част от юрския период и в креда. Примезиофитите достигат максималното си разнообразие през късната креда,
Отдел криптофитни водорасли (cryptomonas) - cryptophyta
Отделът носи името на рода Cryptomonas (от гръцки kryptos - скрит, monas - индивид). Включва едноклетъчни, подвижни, монадични организми. Криптофитни клетки
А Б В Г Д
Ориз. 53. Външен видмогат да образуват криптофитни водорасли (по: G.A. Belyakova et al., 2006): A - Rhodomonas, B - Chroomonas, C - Cryptomonas, D - Hylomonas, E - Goniomonas
Отдел Зелени водорасли - хлорофити
Зелените водорасли са най-обширните от всички отдели на водорасли, наброяващи, според различни оценки, от 4 до 13 - 20 хиляди вида. Всички те имат зелен цвят на талиите, което се дължи на преобладаването на хлоро
Разред Ulotrix - Ulotrichales
Род Ulotrix (фиг. 54). Видовете Ulotrix живеят по-често в сладка вода, по-рядко в морски, солени водоеми и в почвата. Те се прикрепят към подводни предмети, образувайки яркозелени храсти.
Разред бриопсидни – Bryopsidales
Повечето видове се срещат в сладки и солени води. Някои от тях растат върху почва, скали, пясък и понякога солени блата. Род Briopsis - нишковидни храсти до 6-8 s
Разред Волвокс - Volvocales
Родът Chlamydomonas (фиг. 57) включва над 500 вида едноклетъчни водорасли, които живеят в пресни, плитки, добре затоплени и замърсени водоеми: езера, локви, канавки и др. и т.н
Отдел Харофити (Characeae) - Charophyta
Харофити - ред сладководни зелени водорасли, довели до висши растения. Това са форми предимно с нишковиден талус. Често талусът е вертикален, разчленен и носещ се
Отдел динофити (динофлагелати) - dinophyta
1. Името на катедрата идва от гръцки. dineo - завъртане. Комбинира предимно едноклетъчни монади, по-рядко кокоидни, амебоидни или палмелоидни, понякога колониални
Отдел euglenozoa - euglenoids
Разделението на наименованието според типа род е Euglena (от гръцки eu - добре развито, glene - зеница, око). Комбинира единични монадични или амебоидни представители. Рядко се среща в
Речник на термините
Автогамия - полово размножаване, при което две сестрински хаплоидни ядра се сливат в обща цитоплазма. Автоспората е структура на безполово размножаване, което е
Всички хетероконти изглежда имат флагеларен апарат. Камшичетата са 2, като на единия от тях има много характерни тръбести тричленни перести израстъци, или власинки - мастигонеми. Наличието на две неравни флагели послужи като основа за наименуването на тази огромна група водни организми многофлагелативодорасли, или хетероконтами. В допълнение към разликите в дължината, камшичетата се различават и морфологично: основният флагел се състои от ос и ресничести косми, разположени на перисти върху него, страничният флагел е гладък и флагелат. Съставът на пигментите също е подобен - хлорофили АИ си множество каротеноиди, както и структурата на хлоропластите. Хлоропластите са заобиколени от четири мембрани, като двете външни мембрани са свързани с ендоплазмения ретикулум на клетката.
Като част от отдела за водорасли Heterocont (Diversified), няколко групи водорасли са обединени в ранг на класове: кафяви, златни, жълто-зелени, Rafidophytes, Sinura, Feotamnia, Eustigma, някои от които преди са били считани за независими отдели.
Клас Кафяви водорасли– Phaeophyceae (Fucophyceae)
име на клас " кафяви водорасли"отразява цвета на техния талус (от гръцки. Рhaios- мургав). Цветът се дължи на наличието на голям брой кафяви и жълти пигменти - в допълнение към хлорофилите АИ сИ β -каротин в техните хлоропласти има излишък от ксантофили, особено фукоксантин.
Многоклетъчни, предимно големи водорасли, особено широко разпространени в студените води на двете полукълба. Сред кафявите водорасли не са известни нито едноклетъчни, нито колониални форми. Известни са около 1,8 хиляди вида.
Клетъчна структура. Клетката при видовете кафяви водорасли е еукариотна: има образувани органели със собствени мембрани: ядро, митохондрии, пластиди и др. Зелени пигменти - хлорофили АИ с- маскирани от допълнителни: оранжеви каротини и жълто-кафяви ксантофили; от последните преобладава фукоксантинът. В клетките на кафявите водорасли се откриват един до много пластиди. Върху външната клетъчна мембрана клетките на кафявите водорасли обикновено развиват твърда клетъчна стена.
Ядрото най-често е едно, но понякога има няколко ядра. При някои представители с увеличаване на възрастта на клетката броят на ядрата се увеличава. В ядрото винаги има едно ядро.
Хлоропластите на вегетативните клетки на кафявите водорасли имат око (стигма) и пиреноиди, които могат да бъдат един или повече; някои видове нямат пиреноиди. Оцелусът, състоящ се от 30-40 липидни глобули, функционира като леща, която фокусира светлината върху флагеларното издуване, което е фоторецептор. Зооспорите и гаметите на кафявите водорасли обикновено нямат око. Резервните вещества се отлагат в пиреноидите. Хлоропластите най-често са малки, дисковидни, париетални. Тяхната форма може да бъде звездовидна, лентовидна или ламеларна. Формата на хлоропластите в клетките на кафявите водорасли може да се промени с остаряването на клетката. Обвивката на хлоропласта се състои от четири мембрани, като двете външни мембрани са свързани с ER. - ендоплазмения ретикулумклетки.
Целулозата, която изгражда клетъчната стена на кафявите водорасли - понякога наричана алгулоза - се различава по свойства от целулозата на висшите растения. Извън целулозната стена има силно слузест пектинов слой, образуван главно от алгинова киселина и нейните соли и съединения с протеинови вещества. Алгинова киселинасъщо се среща само в кафявите водорасли.
Основни характеристики.
Всички кафяви водорасли са фототрофи. Продукти на фотосинтезата - специфичен въглехидрат хризоламинарин,алкохол манитолИ липиди.Липидите се натрупват под формата на маслени капчици. Хризоламинаринът се отлага извън хлоропласта: той се намира в специални вакуоли в цитоплазмата близо до пиреноида. Манитолът изпълнява резервна, както и осморегулаторна функция; концентрацията му в клетката зависи от промените в солеността на водата.
Младите клетки на кафявите водорасли съдържат малки и многобройни вакуоли, които стават по-големи с възрастта поради сливането. В цитоплазмата има специални образувания – физоди, които съдържат флоротанини.Тези вещества са известни само при видовете кафяви водорасли. Функцията на флоротанините може да бъде свързана със защитата на талиите на водораслите от изяждане от животни, като мекотели. Флоротанините на кафявите водорасли вероятно инхибират заселването на епифитни водорасли и животни на повърхността на талуса. Предполага се, че флоротанините участват в защитата срещу радиационно увреждане и в селективното усвояване от водна средайони на тежки метали. Безцветните флоротанини се окисляват във въздуха, за да образуват пигмента фикофеин, който придава характерния тъмен цвят на изсушените кафяви водорасли.
Структурата на талуса.Сред видовете кафяви водорасли няма едноклетъчни и колониални форми, всички индивиди са многоклетъчни. Формата на талиите на кафявите водорасли е най-разнообразна. Това са нишки, пълзящи или вертикално стоящи, едноредови или многоредови; масивни твърди или керемидени корички и корички; тънки лигавични плочи или плътни кожени плочи на стъблото, прости или разчленени; мехурчета и торбички; тръби и въжета; различно разклонени тънки храсти или мощни храсти. Закрепването на талиите се извършва с помощта на ризоиди или подметки. За да ги поддържат във вертикално положение, големите бентосни форми на кафяви водорасли образуват въздушни (плуващи) мехурчета, пълни с газ (фиг. 29).
Ориз. 29. Външен вид на кафяви водорасли: 1 - Fucus; 2 - Захарин; 3 - Алария; 4 - Саргасум
Таломите на представителите на разредите Laminaria и Fucus са най-сложните сред видовете кафяви водорасли. Те имат признаци на тъканна диференциация със специализация на клетките на талуса (фиг. 30). В техните тали се различават корасъстоящ се от интензивно оцветени клетки и сърцевина, състоящ се от безцветни клетки, които често са събрани във влакна. При Laminaria в сърцевината се образуват и ситовидни тръбички и тръбести нишки. Ядрото изпълнява не само транспортна функция, но и механична, тъй като съдържа нишки с дебели надлъжни стени. Между кората и сърцевината на много кафяви водорасли може да има междинен слой от големи безцветни клетки.
Височинаталусът при представителите на кафявите водорасли е най-често интеркаларен (интеркаларен) и апикален (апикална), рядко базална. Интеркаларният растеж може да бъде дифузен или може да има зона на растеж. При големите форми интеркаларната (интеркаларна) меристема или активно деляща се група клетки се намира в точката на прехода на "петурата" в "листното" острие. Големите водорасли също имат меристематична зона на повърхността на талуса, така наречената меристодерма (вид аналог на камбия на висшите растения).
размножаванев кафявите водорасли, вегетативни, безполови и сексуални. Гаметите се образуват в многоклетъчни гаметангии, зооспорите - в 
Ориз. 30. Вътрешно устройство Ламинария(по: L.L. Velikanov et al., 1991): А- надлъжен разрез на стъблото; б- напречен разрез на стъблото; IN- срез от плоча със соруси на зооспорангии
едноместни спорангии.
Вегетативното размножаване при редица видове кафяви водорасли може да се извърши от местата на талуса; във видовете мил Сфацелария- плодови клонки (плодови пъпки); при видове от рода Фукус- група клетки на подметката, които са способни да се диференцират в нов талус.
Безполовото размножаване на кафявите водорасли обикновено става с помощта на зооспори. Някои видове се размножават чрез неподвижни тетраспори или моноспори. Спорите на асексуалното размножаване се образуват в резултат на мейоза и последващи митози в едноместни спорангии. Половият процес е изогамия, хетерогамия и оогамия. Гаметите се образуват в полиедъртези гаметангии- специални групи клетки. Във всяко „гнездо“ на гаметангиума се образува по една гамета.
За кафяви водорасли, полови феромони- разтворими вещества, които координират дейността на клетките по време на възпроизводството. Феромоните, от които са известни поне десет, могат или да стимулират отварянето на антеридии, или да привлекат мъжки гамети към женски. Те също участват в изолацията на видовете. Смята се, че феромоните се образуват в повечето видове кафяви водорасли, които имат полов процес.
Жизнен цикъл.Кафявите водорасли имат два основни вида жизнен цикъл. Един вид е хаплодиплобионт. Спорофитите са диплоидни, т.е. Всички клетки съдържат пълен набор от хромозоми. Спорите на асексуалното размножаване се образуват в едноместни спорангии. По време на образуването на спорите настъпва редукционно делене - мейоза. Такова намаление се нарича спорно. Образуваните след мейоза клетки (спори) са хаплоидни; съдържа половината от броя на хромозомите. Хаплоидните зооспори и тетраспори покълват в хаплоиден гаметофит, върху който се образуват гамети в многоклетъчни гаметангии. Диплоидната зигота, образувана след сливането на гамети, прераства в диплоиден спорофит.
Друг тип е диплобионт; спорофитите, спорите и гаметофитите са диплоидни; редукционно делене възниква по време на образуването на гамети. Такова намаление се нарича гаметично. След сливането на хаплоидни гамети се образува диплоидна зигота, която покълва в диплоиден спорофит. Този тип е често срещан при видовете водорасли (фиг. 31).

Ориз. 31. Жизнен цикъл Захарини(по: L.L. Velikanov et al., 1991): 1 – спорофит, 2 – соруси на зооспорангии, 3 – зооспори , 4 - гаметофит, 5 - антеридий, 6 - оогониум, 7 - антерозоид, 8 - зигота (оплодено яйце)
На макроскопичен спорофит Захарини, в горната част на плочата се развиват ивици сори(групи) от зооспорангии. В зооспорангиите след редукционно делене се образуват зооспори с две неравни камшичета. Те растат в микроскопични гаметофити: мъжки с антеридии и женски с оогонии. Яйцето, оплодено от антерозоиди, веднага, без период на сън, покълва и образува спорофит. В този жизнен цикъл Захаринизавършва.
Водораслите Fucus нямат спорофити. При Фукусамъжките и женските полови органи - антеридии със сперматозоиди и концептакули с оогонии и яйца - се намират във вдлъбнатините на талуса - скафиди, мъжките и женските скафидии могат да бъдат на едно и също съдили в различни рецепти. Oogonia седи на клетка-крак , появяващи се на стената концептула(фиг. 32, C, D).Оогониевата клетка претърпява три деления

Ориз. 32. Размножаване Фукуса(според: B. Fott, 1971; W. Braune et al., 1974): А – фрагмент от талус (1 – въздушно мехурче, 2 - рецепта); б– разрез през съдовете; IN– разрез на женска скафидия; Ж- оогония; д- освобождаване на яйца; д– разрез на мъжки скафид; И– клон с антеридии; 3 – продукция на сперма
ядра, като първото от тях е редукция, в резултат на което в оогония се образуват 8 яйца.
Антеридиите обикновено се образуват върху разклонени нишки, които ги поддържат - парафизи(фиг. 32, Д,И).В резултат на няколко деления се образуват 64 сперматозоида. сперматозоиди Фукусаимат око, разположено вътре в редуциран хлоропласт. В предния край на сперматозоида се образуват 13 микротубула скитниктекущ- специална структура, която трябва да участва в дефинирането на женския полов атрактант.
Сперматозоидите се привличат към яйцето Фукуса fucoserraten, неспецифичен феромон, секретиран от яйцето. Разпознаването на видово ниво е свързано със специфични олигозахариди в яйцето и спермата. На повърхността на яйцето има различни области, всяка от които съдържа специални органични съединения - гликопротеини. Сперматозоидите съдържат гликопротеини, организирани в места върху мембраната и израстъците на предния камшик, както и на повърхността на самия сперматозоид. При среща с яйцеклетката сперматозоидите се движат по черупката на яйцеклетката, сякаш я изследват с помощта на предния си флагел. Ако гликопротеините на сперматозоида са комплементарни на гликопротеините на яйцеклетката, тогава настъпва само разпознаване на сперматозоида от яйцеклетката.
Систематика
Фосилни находки, които може да са свързани с кафяви водорасли, са известни от средния силур (около 425 Ma). Но тези находки също са подобни на някои съвременни зелени и червени водорасли. Фосилни находки, които определено могат да бъдат свързани със съвременните кафяви водорасли, датират от миоцена (5–25 милиона години). Тези водорасли напомняща на съвременната Dictiota. Възрастта на кафявите водорасли, определена чрез молекулярни методи, е 155–200 милиона години.
Типът организация на талуса, наличието или отсъствието на пиреноид, начинът на растеж, типът на сексуалното размножаване (изогамия, хетерогамия, оогамия) и жизненият цикъл се използват за разграничаване на разредите на кафявите водорасли. През последните години системата на кафявите водорасли беше активно преразгледана. В различни класификационни схеми, като част от клас Кафяви водорасли, се разграничават от 7 до 14 разреда.
Десмарестовски орден–Desmarestiales. Водораслите са храстовидни, многоразклонени, с щипка, плосък или сплескан талус. Захваща се за основата с добре развита подметка. На повърхността на талуса се развиват косми.
Род Десмарестия (фиг. 33). Кафяви или маслинови храсти с добре очертана главна издънка, до 0,8–1 m височина. Прикрепени с конусовидна подметка. Разклонение 3-4 реда, редуващи се или срещуположни. Клоните на последните разреди са плоски или по-рядко нишковидни. Видовете са морски, живеят в арктическите и умерените ширини на дълбочина 1–20 m, образуват гъсталаци или растат поединично.

Ориз. 33. Десмарестия (р. Десмарестия)
Ралфсов орден–Ralfsiales. Представителите на разреда имат талус под формата на корички различна структураили се състои от основна кора и вертикални издънки.
Род Аналипус.Талусът се състои от многогодишна коровидна основна част и от нея излизат едногодишни маслиненокафяви вертикални издънки с височина до 40 см. Многоклетъчните спорангии са с цилиндрична форма и двуредово устройство. Едноместните спорангии са овални и се развиват сред асимилационните разклонения. Видовете от рода са широко разпространени в моретата на умерените ширини, образувайки пояси в приливната зона.
Поръчайте Dictyota–Dictyotales.За видовете от разреда е характерен апикален растеж. Вегетативните клетки съдържат множество дисковидни хлоропласти без пиреноиди. Характерно е паренхимен талус, изоморфно редуване на поколенията. Половият процес е оогамен, безполово размножаване чрез неподвижни спори, сперматозоидите имат само един флагел. Камшикът е дълъг, с перести израстъци и насочен напред. Представителите на разреда се срещат предимно в тропическите и по-рядко в умерените морета.
Род Dictyota (Приложение, 4A). Има талус под формата на плоски маслиненокафяви дихотомично разклонени храсти с височина до 11–15 см. Клоните са линейни, тесни, без жилки, с разклонени върхове. Тетраспорангиите образуват соруси на повърхността на талуса, които имат неопределена или пръстеновидна форма. Разпространен в морските води на умерени и топли ширини.
Поръчайте Ламинария–Laminariales. Ламинарията има талус, състоящ се от плоча, стъбло и прикрепващи органи - подметка или ризоиди. Плочата е равномерна или набръчкана, цяла или разсечена, с дупки, гънки, шишки или без тях, дълга от няколко десетки сантиметра до 20–40 м. Стъблото е разклонено или по-често неразклонено. Спорофитите са едногодишни или многогодишни, някои видове са на възраст 11–18 години. Слизестите канали са разположени в кората на много водорасли.
Най-популярните представители на кафявите водорасли са келп (видове род Ламинария или захарина), или „водорасли“, които са разпространени предимно в Тихи океан. Те се намират от повърхността (ръба) на водата до дълбочина 200 м. Ламинарията расте за предпочитане на дълбочина 2–20 (30) м, на места с постоянно движение на водата, прикрепени към камъни и скали. Ламинарията образува водораслови пояси по крайбрежието на моретата на Далечния изток. Най-мощните гъсталаци се намират близо до Курилските острови и на около. Хокайдо, където са концентрирани около половината от световните ресурси на ламинария. Годишната световна реколта от водорасли водорасли достига 2 милиона тона прясно тегло, докато повече от 1 милион тона идват от производството на марикултура в Китай.
Род Захарин (ламинария).Талусът е дълъг от 0,5 до 5–10 m, има дебела лъскава пластина, еластично стъбло и плътно преплетени ризоиди, които са прикрепени към различни твърди субстрати. Ръбовете на плочата са гладки или вълнообразни, често има издутини и гънки по повърхността на плочата. От двете страни на плочата се образуват натрупвания на спорангии - соруси, след освобождаването на спорите горната част на плочата се унищожава. Живее от две до няколко години. Захарините са ценна промишлена суровина и обект на марикултурата. захарина японскаобразува гъсталаци в северната половина на Японско море, както и близо до южното и югоизточното крайбрежие на Сахалин и Южните Курилски острови. захарина японскае най-ценният търговски вид, има отлични вкусови качества и се използва за приготвяне на различни ястия, консерви, сладкарски изделия.
Разред Sphacelariae–Sphacelariales.
Кафявите водорасли, принадлежащи към този разред, имат разклонен талус с апикален растеж. Космати клони. Вегетативните клетки имат множество хлоропласти без пиреноиди. Жизненият цикъл на мнозинството с изоморфна смяна на поколенията, сексуалният процес е изогамия или хетерогамия. Водораслите образуват малки кафяви гроздове до 3 см височина и разклонени храсти до 20 см височина върху камъни или други водорасли; има ендофитни представители.
Род Сфацелария. То има тънък талус. Разклонените нишки образуват малки, до 0,5–3 cm високи, кафяви снопчета. Вегетативното размножаване се извършва от специализирани, често трилъчеви разклонения - пропагули. Половият процес е изогамен. Женските гамети секретират феромоните ектокарпен и мултифиден. Видовете от рода са разпространени в моретата както на полярните, така и на тропическите ширини.
Разред Сцитосифон–Scytosiphonales. Представители с чревен, мехурчест, пластинчат или торбовиден, често кух талус с дължина до 25 cm. Кората е едноредова или двуредова, вътрешната част (стена) на талуса е образувана от няколко слоя големи клетки. На повърхността на талуса се развиват многоклетъчни спорангии, които са събрани в соруси, които поддържат техните нишки - парафизии снопчета косми.
Род Сцитосифоне тънкостенна кафява тръба с дължина 4–25 cm (Приложение 4B). Талусът е прикрепен с малка подметка и има редки напречни стеснения. Среща се в крайбрежната зона на моретата на различни географски ширини, често образува пояси в приливната зона.
Фукус ред–Фукалес. Имат многогодишен талус под формата на големи неравномерно или дихотомно разклонени храсти с дължина до 1–3 m. Те са прикрепени към субстрата с конусовидна подметка. Талусите имат издънки, филоиди (листовидни образувания), вместилища (вместилища на гамети) и плувни (въздушни) мехури. Те се срещат в приливната зона на всички умерени и арктически морета, където образуват мощни гъсталаци - пояси.
Род саргасъм(фиг. 34, IN). Обилно разклонени маслиненокафяви или кафяви храсти с височина 0,5–1,5 м. Прикрепени с конична подметка. Филоидите се развиват върху странични издънки, излизащи от цилиндричното стъбло в долната и средната част на талуса. Филоидите са кожести, с централно ребро, с дължина 0,2–2 cm. Плувните мехури са овални или заоблени, със или без израстъци. Sargassums са широко разпространени в умерените и субтропичните ширини от долното крайбрежие до дълбочина 10 m, където образуват гъсти или редки гъсталаци. В южната част на Тихия океан - така нареченото Саргасово море - те образуват обширни плаващи натрупвания, в които се размножават само вегетативно.
Род Фукус (фиг. 32).Талус кожен, храстовиден, разклонен дихотомно, висок 25–50 см. В близост до брега на далекоизточните морета расте няколко години, живее в крайбрежната зона. Често образува големи гъсталаци в плитки води. Видовете от този род се характеризират с дихотомия
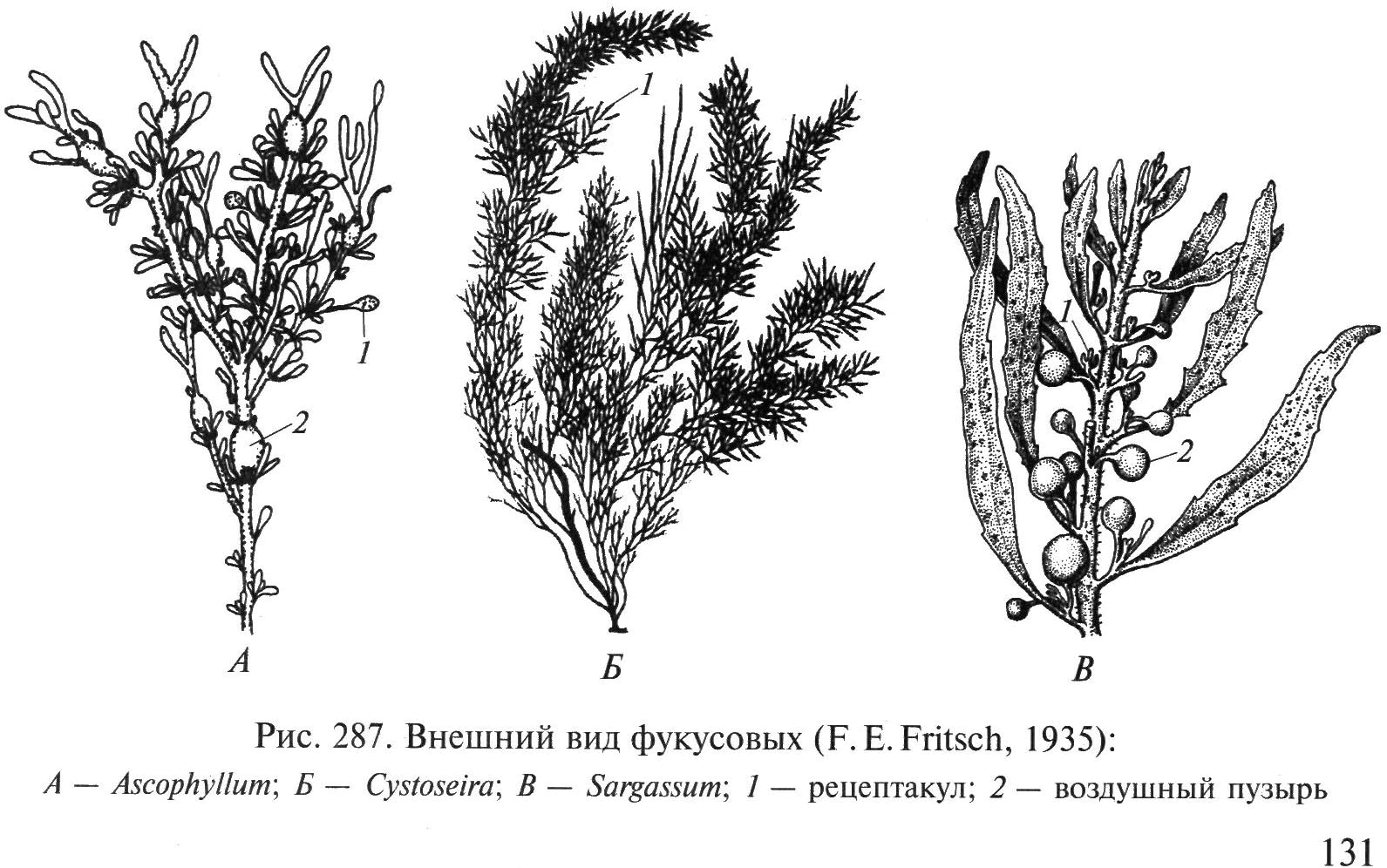
Ориз. 34. Появата на водорасли фукус: А – Аскофилум; б – Cystoseira; IN – саргасъм; 1 - съд, 2 - плувен мехур
разклонен талус с плоски клони с надлъжно ребро, който е прикрепен към камъните с конична подметка. Фукусите се използват като торове, като храна за добитък, за производство на фуражно брашно, алгинати, използвани в медицината за лечение на гуша, корекция на теглото и др.
Разред Ectocarps–Ectocarpales. Водораслите, които са включени тук, са микроскопични, свободно живеещи, епифити, епизоити или ендофити. Репродуктивните органи са представени от едноместни (спорангии) и многоместни (гаметангии) спорангии; биват дългоцилиндрични, късоцилиндрични или шушулковидни.
Род ектокарпус. Талус под формата на меки кафяви храсти до 2 см височина. Храстите са оформени от едноредово, последователно, дихотомно или неправилно разклонени нишки. Многоклетъчните спорангии са шушуловидни, образувани отстрани на страничните разклонения на 1–3-клетъчна дръжка. Видовете са разпространени в моретата на различни географски ширини, в чисти и замърсени води, в литорала и в горната част на сублиторала. Епифитите се срещат и в обрастването на антропогенни субстрати.
Екология
По-голямата част от кафявите водорасли живеят в морските води. В сладките води се срещат само 8 вида. Има едногодишни и многогодишни видове, чиято възраст може да достигне 15-18 години. Ламинария водорасли НереоцистисИ Макроцистис, които живеят по тихоокеанското крайбрежие на Северна Америка, имат максималния размер за всички водорасли - до 30–40 m дължина. Те образуват гигантски подводни гори в моретата на Далечния изток. Кафявите водорасли се прикрепят към голямо разнообразие от субстрати - скали, камъни, чакъл, раковини и раковини на морски животни и други водорасли. Някои малки форми на кафяви водорасли живеят в тъканите на други водорасли като ендофити. Повечето кафяви видове живеят в прикрепено състояние. Откъснатият от земята талус се отнася от течението до спокойни места с тинесто дъно и продължава да съществува там. Видовете с въздушни мехурчета върху талуса, откъсвайки се от земята, изплуват на повърхността, образувайки големи плаващи клъстери, особено в райони със стабилно кръгово течение, като в Саргасово море. Те съществуват в такива клъстери доста дълго време и се размножават само вегетативно.
Кафявите водорасли са широко разпространени във всички морета, но най-голямо развитие достигат в моретата на умерените и субполярните ширини. Техните големи тали растат прикрепени към скали и камъни, а на тихи места близо до брега и на големи дълбочини те дори могат да се държат върху клапите на черупките на мекотели и чакъла. Можете да ги срещнете на различни дълбочини - от крайбрежната зона, където при отлив те са извън водата с часове, до 40–200 м. В района на Хавайските острови, на дълбочина 180 м, видове от рода саргасъм, А Келп V Адриатическо море е открито на дълбочина 200 м. Най-изобилните гъсталаци от кафяви водорасли се наблюдават на дълбочина 6–15 м, където най-добрите условия на осветление и постоянното движение на водата, което носи хранителни вещества на техните тали и граници заселване на тревопасни животни - фитофаги.
Клас Златни водорасли -Chrysophyceae
Едноклетъчни, колониални или многоклетъчни организми, обикновено имащи флагеларен стадий в своя жизнен цикъл. Клетките са едноядрени, с един или повече хлоропласти. Обикновено има стигма (фиг. 36). Камшичетата 1–2, неравни. Съкратителната вакуола е разположена в предната част на клетката. Външните обвивки на клетките на тези организми са много разнообразни. При най-примитивните видове клетката е облечена в деликатен перипласт, който позволява образуването на издатини с различна форма (ризоподии, псевдоподии), при други плазмалемата е покрита с твърда целулозна мембрана, понякога силно слузеста. Къщите в златните водорасли се предлагат в различни форми: във формата на ваза, сферична, яйцевидна, цилиндрична; с една или повече дупки. Клетката е прикрепена към основата на къщата с гъвкав крак или може да бъде свободна. При някои хризофити върху клетъчната повърхност се отлагат варовити образувания - коколити - с различни форми и размери, а силициевите флагелати имат вътрешен силициев скелет. Разнообразието на структурата на тези скелети дава възможност да се използват останките от различни форми на изкопаеми силикофлагелати за определяне на възрастта на геоложките скали, които ги съдържат. При по-силно развитите представители на отдела на златните водорасли клетките са облечени в черупка, състояща се от силициеви люспи (понякога носещи шипове) или затворени в къщи, през отворите на които излизат флагели или псевдоподии.
Къщите при видовете златни водорасли са във формата на ваза, сферични, яйцевидни, цилиндрични, с една или повече дупки. Клетката е прикрепена към основата на къщата с гъвкав крак или може да бъде свободна.
Пигменти - хлорофили АИ с, фукоксантин. Хризофитите се характеризират с най-високо съдържание на фукоксантин сред хетероконтите (55-93% от всички каротеноиди).
Повечето видове са сладководни фототрофи, но се срещат и хетеротрофи и фаготрофи.
Резервни вещества - хризоламинарин и масла.
Сред едноклетъчните форми на златните водорасли преобладават сладководни планктонни видове, а сред многоклетъчните форми - бентосни или епифитни. Срещат се главно в хладни морски и сладки водоеми. Известни са около 360 вида.

Ориз. 36. Златни водорасли (по: C. Hoek van den et al., 1995): А- клетъчна структура; б- коренова система; IN- трипартитен мастигонема; Ж- разрез на част от хлоропласта; 1 - базално тяло; 2 - базално подуване; 3 - къс флагел; 4 - стигма; 5 - ризопласт; 6 - ядро; 7 - ядрена мембрана; 8 - ядро; 9 - митохондрия; 10 - вакуоли; 11 - слуз; 12 - плазмалема; 13 - везикули; 14 - лигавично тяло; 15 - липиди; 16 - хлоропласт; 17 - апарат на Голджи; 18 - контрактилна вакуола; 19 - дълъг камшик: 20 - 1-ви корен; 21 - 2-ри гръбнак; 22 - 3-ти гръбнак; 23 - 4-ти корен; 24 - хлоропластен нуклеоид; 25 - HES; 26 - хлоропластна обвивка; 27 - поясна ламела; 28 - ламела; 29 - дълга странична нишка; 30 - къса странична нишка; 31 - крайна нишка; 32 - тръбна част на мастигонема
структура на талусав златните водорасли най-често е монадичен, но може да бъде много разнообразен: кокоиден, палмелоиден, плазмоиден, амебоиден, нишковиден, паренхим (фиг. 37).

Ориз. 37. Златни водорасли (по: L.L. Velikanov et al., 1981): А – Охромонас: 1 - външен вид, 2 - киста; б – Хромулина: 1 - външен вид, 2 - киста, 3 - схема на образуване на филм от кисти; IN – Динобрион: 1 - общ изглед на колонията, 2 - киста; Ж – Хризамеба; д – гибердия; д – Хидрурус: 1 - общ изглед на колонията, 2 - връх на клона, 3 - зооспора
размножаване. При златните водорасли има три начина на размножаване: вегетативен, асексуален и сексуален.
Вегетативно размножаваневъзниква чрез надлъжно клетъчно делене или разпадане на колония на части поради клетъчно делене в една, две или три посоки.
полов процес(клетките на златните водорасли са диплоидни) - изогамия или конюгация. В резултат на половия процес, както и при неблагоприятни условия се образуват кисти с плътна обвивка, съдържаща силиций.
безполово размножаванеизвършва се от едно- или двуфлагелирани зооспори, които се развиват в обикновени вегетативни клетки или зооспорангии. Сексуален процес: холо-, изо- и автогамия. Хологомията е вид полов процес, при който две клетки, подобни на вегетативните клетки, се сливат в предните краища в двуядрена зигота, която след това се превръща в силикатизирана киста.
Систематика
От златните водорасли в края на миналия век въз основа на резултатите от молекулярно-генетични изследвания е изолиран клас Sinuraceae. Разграничава се и клас Feotamniaceae - от жълтозелени и златни водорасли. Поради това обхватът на клас Chrysophyceae е намален и в момента включва около 360 вида, разпределени в три разреда: Chromulinales, Hibberdiales и Hydrurales. Формата, структурата и броят на камшичетата, както и пигментният състав и структурата на тялото са важни систематични характеристики.
Разред хромулин - Chromulinales. Този ред включва организми с монаден, палмелоиден и амебоиден тип диференциация на талуса. Монадични клетки с един флагел, видим под светлинен микроскоп.
Род Хризамеба(фиг. 37, А) - сладководни амебоидни водорасли, те имат флагеларен стадий в жизнения си цикъл. Зиготата се развива в стоматоциста.
Род Хромулина(фиг. 37, IN) - едноклетъчни свободно плуващи организми, които живеят предимно в сладки води. Клетките са голи; има един или два златни пластида.
Род Динобрион(Фиг. 37 , G)- едноклетъчни и колониални, свободно плаващи или прикрепени представители. Монадите са разположени в къщи с форма на ваза, образувани от целулозни микрофибрили. В допълнение към целулозата, къщата съдържа и голям брой аминокиселини. Храстовата форма на колониите е свързана с метода на вегетативно клетъчно делене, когато една от дъщерните клетки, напускайки родителската къща, се прикрепя към дупката си. Въртейки се около оста си, тя образува собствена къща. И двете дъщерни клетки могат също да напуснат родителската къща. Клетките съдържат 1-2 хлоропласта; в предния край на клетката се виждат око и 2 контрактилни вакуоли. Хризоламинаринът се намира във вакуола в задния край на клетката. Размножаването е вегетативно и полово.
Разред Gibberdiae–Hibberdiales. Представителите на този ред се характеризират с уникален пигментен състав. В допълнение към фукоксантина, те имат допълнителен светлосъбиращ каротеноиден пигмент, антераксантин.
Включва раждане Gibberdia, Chromophyton, Styloceras, Chrysopixis, Platytekaи други.
Род гибердияима два етапа в жизнения цикъл: колониален палмелоиден неподвижен и едноклетъчен монаден подвижен. При светлинна микроскопия се вижда само един флагел в монадния стадий. В допълнение към фукоксантина, характерно е наличието на друг допълнителен светлосъбиращ каротеноиден пигмент, антераксантин.
Поръчайте Hydrorus–Hydrarales. Представителите на този ред имат талус с палмелоиден и псевдопаренхимен тип структура. Характерно е наличието на уникални тетраедрични зооспори. Няма око. Митозата е полузатворена.
Включва раждане Hydrorus, Celloniella, Federmatium, Chrysonebulaи други.
Род Хидрурус(фиг. 37, д) е най-диференцираният сред палмелоидните водорасли. Талусът му представлява големи, до 30 см дълги лигави колонии, които приличат на кафяви разклонени нишки, често излъчващи много неприятна миризма. В колониите може да се направи разлика между главния ствол и страничните клони. Клетките, потопени в обикновена слуз, съдържат един чашковиден хроматофор и няколко контрактилни вакуоли. Клетките са рехаво разположени по периферията на колонията и по-плътни в средата. Талусът може да расте само чрез делене на апикални клетки. При безполово размножаване в клетките на страничните разклонения на колонията се образуват зооспори. Образува сферични кисти. Среща се в планински потоци и реки със студена вода, където се прикрепя към твърди субстрати.
Клас Синура водорасли -Sinurophyceae
син- заедно и ура- опашка. Тази група организми е изолирана през 1987 г. от златни водорасли. Той обединява монадични единични и колониални организми, понякога с редуване в жизнения цикъл на монадични и палмелоидни стадии. Повърхността на пектиновите черупки е покрита с обвивка от силициеви люспи. Митохондриите с тръбести кристи обикновено са разположени в цитоплазмата около хлоропласта. Ядрото е едно, хлоропластите са заобиколени от четири мембрани, обикновено две. Ламелите са тритилакоидни, има поясна ламела. Основните пигменти са хлорофилите АИ с, β -каротин и фукоксантин. Няма стигма. Клетките обикновено с 2 неравни флагела. Дългият перест камшик е насочен напред. Къс, гладък камшик, понякога силно намален, насочен назад.
Клетките се размножават главно чрез надлъжно делене. Колониите, разпадайки се, пораждат млади колонии. При някои видове половият процес е описан под формата на изогамия. В същото време, за разлика от златните водорасли, сливането на изогамети в синуричните водорасли се случва не в предните, а в техните задни краища. Силицираните цисти с пори се образуват ендогенно, подобно на тези на хризофитите.
Преобладаващо планктонни форми, палмелоидни стадии са част от бентоса. Повечето видове синурови водорасли са фототрофни сладководни организми.
Систематика.
Класът Sinura водорасли включва 7 разреда: Chloramoebales, Synurales, Rhizochloridales, Ochromonadales, Heterogloeales, Parmales и Thallochrysidales.
Разред Синура - Synurales. Монадичните форми, пектиновите клетъчни мембрани обикновено са покрити с черупка от силициеви люспи, циментирани с органична материя в един корпус. Хлоропласти един или два.
Род Синура(фиг. 38) - сладководни монадични форми с два неравни флагела и два париетални хлоропласта. Ядрото е с крушовидна форма, разположено в предната част на клетката. Зад ядрото има една голяма вакуола, а в задната част на клетката има няколко малки контрактилни вакуоли. Клетките са покрити с люспи, като плочки.Размножаването е най-често вегетативно, при някои е познат половият процес. Образува колонии (фиг. 39, А). Често кистите се появяват във всички клетки на колонията. С масово развитие Синураможе да придаде на водата неприятна миризма.

Ориз. 38. Външен вид на клетките на синура
Разред Ризохлоридни - Rhizochloridales. Предимно сладководни организми с ризоподиален тип диференциация на талуса, живеещи предимно в сладки водни тела.
Род Маломонас(фиг. 39, Б-Д) – едноклетъчна монада с един добре изразен флагел и един раздвоен хлоропласт.

Ориз. 39. Външен вид на синурични водорасли (по: G.A. Belyakova et al., 2006): А – деляща се колония Синура; Б, В – образуване на статоспори и Ж – външен вид Маломонас
Клетката е покрита с керемидени и спираловидни люспи, някои от тях носят игловидни шипове (фиг. 39, Ж). Характерно е наличието на статоспори (фиг. 39, Б, В). За редица представители е описан сексуалният процес, хологамия.
Род Миксохлорисживее в клетките на листата на сфагнови мъхове, има вид на голям многоядрен плазмодий. През есента образува цисти, които покълват през пролетта. От тях се появяват зооспори или амеба, които проникват в празни листни клетки и се сливат там в плазмодий.
Разред Chloramoebaceae - Chloramoebales.Включва представители на монадите. Среща се в солени и сладки води.
Род хетерохлорис – клетките му могат да променят формата си, образувайки псевдоподии. Това явление е присъщо на много представители на разреда, както и склонността към животинския начин на хранене. В клетката има няколко хлоропласта, маслени капки и хризоламинарин. Пред клетката има контрактилни вакуоли, в центъра - едно ядро. Размножава се вегетативно чрез клетъчно делене.
Разред Охромонадови - Ochromonadales. Съчетава голи форми с две неравни флагели. Сладководни и морски форми.
Род Охромонасвключва едноклетъчни монади с две неравни флагели. Клетките са облечени само от плазмалема. В предния край на клетката има контрактилна вакуола и око, в задния край има вакуола с хризоламинарин. Размножава се вегетативно, браздата на делене започва в предния край на клетката между две двойки флагеларни основи. Видовете от рода са често срещани в олиготрофните сладки води, но има и морски представители.
Разред Хетероглееви - Heterogloeales. Включва водорасли с палмелоиден тип диференциация на талуса. Представителите на разреда се срещат по-често в сладки, отколкото в солени води.
Род Хелминтоглеяживее в солени води. Тази колония, разположена върху разширена основа, се състои от разклонени лигави ленти. Протопластите са произволно подредени в тази слуз, всеки от които е заобиколен от собствена лигавична обвивка.
Клас Phaeotamnia водорасли - Phaeothamniophyceae
Името на класа произлиза от типа genus Феотамнион(от гръцки. phaeos- мургав храст). Представители на тази група организми бяха идентифицирани в отделен клас от жълто-зелени и златни водорасли през 1998 г. въз основа на анализа на последователността на рибозомните гени и особеностите на клетъчната ултраструктура. Водораслите Feotamnia се характеризират с уникална комбинация от пигменти: фукоксантин с хетероксантин, липса на виолаксантин.
Характеристики на клетъчната структура: липсата на вакуоли с хризоламинарин и липсата на ендогенни кисти със стени от силициев диоксид.
Класът включва едноклетъчни, колониални и многоклетъчни организмис кокоиден, палмелоиден и нишковиден тип структура на талуса. Камшичести стадии с две неравни камшичета. Камшичета странични или субапикални. Стигмата се среща в зооспорите. Пигменти - хлорофили АИ с, β -каротин, фукоксантин, диадиноксантин, диатоксантин и хетероксантин. Основният резервен продукт е парамилон ( β -глюкан). Клетъчната стена присъства, по време на клетъчното делене дъщерната стена се образува вътре в родителската стена.
При feotamniaceae от основните методи на размножаване се срещат само вегетативно и безполово. Безполовото размножаване се осъществява чрез автоспори или зооспори. Сексуалното размножаване не е известно.
Видовете водорасли Feotamnia живеят изключително в сладки води. Систематика.
Понастоящем е потвърден монофилетичният клас на водораслите феотамния, а сред охрофитите той е по-близо до кафявите и жълто-зелените водорасли, отколкото до златните водорасли. В момента са известни около 30 вида от тези водорасли, приписани на един разред Теотамния - Phaeothamniales.
Род Теотамнионпредставлява прикрепени разклонени нишки с височина до 1 cm (фиг. 40). Клетките са цилиндрични, неправилни, разширени нагоре, надлъжно

Ориз. 40. Външен вид Теотамнион.
два до три пъти по-дълъг от широк. По време на клетъчното делене материалът на родителската стена се изразходва за образуването на слоеста обвивка, обграждаща нишковидните клетки. По периферията на клетката под плазмената мембрана има везикули. Наподобяват физоди – образувания, намиращи се в клетките на кафявите водорасли. Клетките съдържат от един до няколко дисковидни маслиненокафяви пластиди. Зооспорите се образуват от 1–2, по-рядко от 4–8 в една клетка. При зооспорите флагелите са прикрепени странично. Положението на флагеларните корени Теотамнионнаподобява тази на жълто-зелените и кафявите водорасли. Липсва му и ризопласт. В жизнения цикъл могат да се образуват кисти, но в стените им няма силиций. Цистите покълват като зооспори. Заселва се като епифит върху нишковидни водорасли. Обитава стоящи и бавно течащи сладки водоеми.
Клас Rafid водорасли -Raphidophyceae
Името на класа идва от гръцки рафид - игла. Обединява едноклетъчни бифлагелатни организми, лишени от клетъчна стена. Предимно сладководни организми със сплескани клетки (фиг. 41).
Клетките имат флагели с различна дължина; камшикът, насочен напред, е перест и по-дълъг, камшикът, насочен назад, е гладък и по-къс. Преходната спирала липсва. Хлоропластите са малки, многобройни или по-рядко 1-2 на клетка, заобиколени от четири мембрани, от които две са CES мембрани. Тилакоидите са подредени в купчини по три. Ядрото е голямо, заобиколено от пръстен от диктиозоми. Митохондрии с тръбести кристи. Има контрактилни вакуоли. Цитоплазмата често е вакуолизирана. Клетките са голи, заобиколени само от плазмалемата. Стигма (око) обикновено отсъства.
Фотосинтетични пигменти – хлорофили АИ с, β -каротин, вошериаксантин, диноксантин, диадиноксантин, хетероксантин.
Резервни продукти за асимилация са мазнини и масла, по-рядко нишесте и хризоламинарин.
Те също живеят в сфагнови блата и други местообитания с кисела и неутрална водна реакция. Някои видове се срещат в солени и морски води.

Ориз. 41. Външен вид на рапидните водорасли.
Систематика.
Дълго време тази група водорасли, поради зеления си цвят, наличието на фаринкс и редица други структурни характеристики, се считаше за клас Chloromonas сред Euglenoids. Но данните от цитологията, биохимията, физиологията и молекулярната биология показаха безспорната им принадлежност към отдела на водораслите Heterocont (Ochrophyte) и монофилетичната природа на групата водорасли Rafid.
От самото са известни около 25 вида разред Huttonellaceae– Chattonellales.
Род хетеросигма(фиг. 42, А) включва морски флагелати. Клетки с овална форма с флагели, изместени към странична страна, пластидите са разположени по периферията на клетката. Няма трихоцисти. Те плуват, извършвайки въртеливи движения около надлъжната ос на тялото. Може да причини токсичен "цъфтеж на водата" в крайбрежните води.

Ориз. 42. Рафидни водорасли (от: R. E. Lee, 1999): А – хетеросигма; б – Гониостомум; 1 - хлоропласт; 2 - мукоцист; 3 - контрактилна вакуола; 4 - трихоцист; 5 - ядро
Род Гониостомум(фиг. 42, б) са подвижни дорзовентрално компресирани монади. Дорзалната страна е извита, коремната страна е сплескана. Една бразда в предния край води в триъгълен фаринкс, от който излизат две дълги камшичета; тяхната дължина е сравнима с дължината на клетката. Пластидите са разположени по периферията на клетката. Под плазмалемата има трихоцисти. Живее в сладки води с кисело pH.
Клас Eustigma водорасли -Eustigmatophyceae
Името на класа идва от гръцки ЕС- "добре и стигма- "белег", "петно". Класът обединява голи едноклетъчни, по-рядко колониални организми с предимно кокоидна структура.
Едно или повече ядра. Хлоропластът обикновено е един, заобиколен от 4 мембрани. Камшичето обикновено е едно предно пересто и второ базално тяло или по-рядко две неравни камшичета. Налице е стигма (око), разположена извън хлоропласта, нейните гранули са без мембрани.

Ориз. 43. Външен вид на водорасли Eustigma
Хлоропластът обикновено е единичен, голям, многолостен, чашовиден или париетален, жълто-зелен на цвят. Тилакоидите са събрани в ламели по три, обграждаща ламела липсва.
Основните пигменти са хлорофил а, β-каротин, виолаксантин и вошериаксантин. Пиреноидът обикновено присъства само във вегетативни форми.
Резервни вещества са масла и вещество с неизвестна природа, което се отлага като твърд материал извън пластидите. Нишестето не се образува.
Те живеят в сладки води, по-рядко се срещат в моретата и в почвата.
Систематика.
Преди това тази група организми е била класифицирана като жълто-зелени водорасли. В момента водораслите Eustigma се считат за ранг на клас в отдела на Heterocontophytes. Известни са около 35 вида, предимно сладководни, приписвани на единствения Разред Еустигма - Eustigmatales.
Род Евстигматос(фиг. 43) - едноклетъчни малки сферични водорасли. Килийната стена е гладка, цяла, без орнаменти. Жълто-зелен хлоропласт, единичен, дялан, теменен, с дръжкови полиедрични пиреноиди. Голяма централна вакуола с червено съдържание. Размножава се от 2 или 4 автоспори или зооспори във формата на бутилка с един входящ камшик, голяма стигма, разположена извън хлоропласта в предния край, и единичен заден хлоропласт без пиреноид. Често срещан компонент на почвената флора. Известно от Нова Зеландия, Австрия, Исландия и от басейните на Аризона като "синапено водорасло".
Клас Жълто-зелени водорасли -Xanthophyceae
Жълто-зелените водорасли са водорасли, чиито хлоропласти са оцветени в светло или тъмно жълто, много рядко зелено и само от време на време синьо. Цветът на талиите се определя от наличието на следните пигменти в клетките на хлоропластите - хлорофили АИ с, β -каротин и ксантофили. Преобладаването на последното определя оригиналността на цвета на жълто-зелените водорасли. В допълнение, парамилон, маслени капки и само при някои видове, в допълнение, бучки от левкозин и волутин се натрупват в клетките като основен продукт на асимилацията. Жълто-зелените водорасли не образуват нишесте. Отличителна черта на жълто-зеленото е наличието на монадична структура във вегетативните клетки и две неравни флагели в зооспорите. Клетъчната стена съдържа целулоза, глюкоза и уронова киселина. Клетъчната стена често се състои от две части.
Размножаването е вегетативно, безполово и полово.
Широко разпространен в сладки води. Рядко се среща в морски, солени води и в почвата.
Преди това класът жълто-зелени водорасли се е наричал водорасли Tribophytium, след вида на рода Tribonema (от гръцки. трибон – умел, хитър и няма – нишка). Известни са около 450 вида.
Жълтозелените се характеризират със значително морфологично разнообразие. Сред многобройните представители на този отдел се срещат почти всички основни типове структура на тялото: амебоидна, монадична, палмелоидна, кокоидна, нишковидна, многонишкова, ламеларна и сифонална (фиг. 44). – 46). Талус едноклетъчен,

Ориз. 44. Външен вид на жълто-зелени водорасли: 1, 2 - Харатиопсис, 3 – Centritractus, 4 – офиоцитиум
колониални, многоклетъчни и неклетъчни. Клетъчната мембрана е плътна, пектинова и целулозна, състояща се от плътно припокриващи се части или от две клапи. В черупката се отлага силициев диоксид или вар. Предимно фиксирани форми. Сред едноклетъчните видове има подвижни форми, лишени от плътна обвивка и оборудвани с флагели, лобоподии и кореноподии.

Ориз. 45. Външен вид на ксантококови жълто-зелени водорасли: 1–3 - Ботридиопсис, 4 – тетраедриела, 5 – Pseudostaurastrum, 6 – Гониохлорис, 7, 8 – Бумилериопсис
Повечето жълто-зелени – неподвижни организми. При подвижните индивиди движението може да се извършва с помощта на жгутици или ризоподии. Клетки с различни форми: сферични, вретеновидни, елипсовидни, цилиндрични, тетраедрични, сърповидни, крушовидни, яйцевидни. Размери на талуса от 0,5 – 1,5 µm ( Хлоридела) до няколко милиметра в диаметър ( Ботридиопсис) (фиг. 45, 1 – 3) и до десетки сантиметри дължина ( Вошерия) (фиг. 46, 3).

Ориз. 46. Външен вид на жълто-зелени водорасли: 1 - Трибонема, 2 – хетеропедия, 3 – Вошерия, част от нишка с оогоний и антеридий
Повечето жълто-зелени видове са фототрофи, но се открива и холозойно хранене чрез поглъщане на бактерии и малки водорасли. Жълто-зелените водорасли са широко разпространени в сладките води. Те също са често срещани в почвата, по-рядко в морските и солените води. Класът включва аеробионтни, планктонни, бентосни и перифитонни форми. Епифити, епизоити, както и вътреклетъчни симбионти в протозойни клетки.
Независимо от външната структура, вътрешната структура на клетката на жълто-зелените водорасли е една и съща. В протопласта обикновено има няколко жълто-зелени хлоропласта, които имат дисковидна, коритовидна, ламеларна, по-рядко лентовидна, звездовидна или чашковидна форма с цели или лобовидни ръбове. Оцветяването се дължи на липсата на фукоксантин, който е отговорен за златния и кафяв цвят при други охрофити. От другите пигменти, които имат β -каротин, вошериаксантин, диатоксантин, диадиноксантин, хетероксантин. При подвижните форми червено око или стигма обикновено се намира в предния край на хлоропласта. Малко видове имат пиреноиди от полупотопен тип. Ядрото в клетката обикновено е едно, малко по размер, но има видове с многоядрени клетки. Някои видове имат една или две контрактилни (пулсиращи) вакуоли в предната част на клетката.
Представителите на монадите и подвижните стадии (зооспори и гамети) имат два неравни флагела. Изключение правят синзооспорите. Вошери, в който по повърхността са разположени многобройни двойки гладки камшичета, леко различни по дължина. Къс флагелум завършва с акронем. Флагелите са прикрепени към клетката субапикално. При сперматозоида Вошеристранично закрепване.
При видовете с амебоидна, монадална и палмелоидна организация клетъчната стена отсъства, те са покрити само от цитоплазмена мембрана и могат лесно да променят формата си. Понякога "голи" клетки се намират вътре в къщи, чиито стени могат да бъдат боядисани в кафяво с манганови и железни соли. По-голямата част от формите имат клетъчна стена, състояща се от две части. Целулозата преобладава в състава на клетъчната стена, а също така съдържа полизахариди, състоящи се главно от глюкоза и уронова киселина. В младите клетки мембраната е тънка, с възрастта се удебелява. В него могат да се отлагат железни соли, чиито съединения го оцветяват в различни нюанси на кафяво и червено. По-често силициевият диоксид присъства в клетъчната стена, което й придава твърдост и блясък. Може също така да бъде инкрустиран с вар и да бъде изваян по различни начини (бодли, клетки, брадавици, черупки, зъбци и др.) Израстък на черупката може да се образува в прикрепени форми – крачол с подметка за прикрепване.
При нишковидните форми на жълто-зелени водорасли с двучерупчести черупки, когато нишките се разпадат, клетъчните мембрани се разпадат на Н-образни фрагменти. Тези фрагменти са плътно свързани половини на черупките на две съседни клетки (фиг. 47). По време на растежа на нишките, Н-образен фрагмент от клетъчната стена на две съседни дъщерни клетки се вгражда между двете половини на мембраната на майчината клетка. В резултат на това всяка от дъщерните клетки е наполовина покрита със старата мембрана на майчината клетка и наполовина – новообразувана черупка.

Ориз. 47. Схема на образуване на напречна преграда между две дъщерни клетки в нишковидни жълто-зелени водорасли (според: A.A. Masyuk, 1993): А- фрагмент от нишка; б- полагане на поясния пръстен и образуване на напречна преграда между две клетки; IN– наслояване на двучерупчести клетъчни мембрани; Ж– разпадане на черупката на Н-образни участъци
Съкратителните вакуоли присъстват в подвижните представители. Обикновено има 1-2 на клетка. Апарат на Голджи със специфична структура. Диктиозомите са малки, съдържат 3-7 цистерни. Ядрото е едно, по-рядко са много; при ценотичните видове клетките винаги са многоядрени.
размножаване. Повечето видове жълто-зелени водорасли се характеризират с вегетативно и безполово размножаване.
Вегетативно размножаванеизвършено различни начини: клетъчно делене наполовина, разпадане на колонии и многоклетъчни тали на части. При Вошериобразуват се специални плодови пъпки.
При безполово размножаванеМогат да се образуват различни спори: амебоиди, зооспори, синзооспори, автоспори, хемизооспори, полуавтоспори, апланоспори. Зооспорите са "голи" и обикновено имат крушовидна форма.
полов процес- изогамия, хетерогамия и оогамия - описани в няколко представителя. При Трибонемигаметите са сходни по размер, но се различават по поведение – това е изогамия. При Вошеринаблюдава се оогамия: върху нишките се образуват вместилища на женски гамети – оогония и мъжки – антеридии.
IN неблагоприятни условиясе наблюдава образуване на кисти. Цистите (статоспорите) са ендогенни, едноядрени, рядко многоядрени. Стената им често съдържа силициев диоксид и се състои от две неравни или по-рядко равни части.
Систематика.
В края на XIX – началото на 20 век различни родове жълто-зелени водорасли бяха причислени към отдела на зелените водорасли, което беше свързано предимно с цвета и морфологичното сходство на талиите. Жълтозелените понастоящем се считат за клас в рамките на охрофитния отдел.
Известни са около 450 бр съвременни видовеклас Жълто-зелени водорасли, които са групирани в четири разреда: Botridia, Mishococcus, Tribonem и Wocheriaceae. Класификацията на разредите се основава на вида на диференциацията на талуса и характеристиките на жизнения цикъл.
Разред Ботридиеви - Botrydiales. Разредът включва видове със сифонален тип диференциация на талуса, при които липсва оогамен полов процес.
Род Ботридиумживее върху почвата и има вид на зелени мехурчета с размери няколко милиметра, прикрепени с безцветни ризоиди. Талусът е сифонален, съдържа множество ядра и пластиди. Черупката е многопластова, върху нея може да се отлага вар. Размножаването е безполово с помощта на бифлагелатни зооспори, докато цялото съдържание на пикочния мехур се разпада на едноядрени фрагменти. При липса на влага се възпроизвежда с помощта на апланоспори или образува дебелостенни кисти. В някои случаи цялото съдържание на пикочния мехур отива до образуването на една голяма киста. В други случаи кистите се образуват в ризоидите, където първо преминава съдържанието на пикочния мехур. Цистите покълват или директно в новия талус, или образуват зооспори. Половият процес е изо- и хетерогамия. Зиготата покълва веднага, без период на покой. Често срещан и широко разпространен вид в сухоземните местообитания, намиращ се по бреговете на потоци, езера или върху почви без растителност.
Поръчайте Michococcal–Mishococcales. Едноклетъчни или колониални представители с кокоиден тип диференциация на талуса.
Род Харатиопсисвключва едноклетъчни прикрепени форми. При размножаване образува зооспори, апланоспори и дебелостенни цисти (фиг. 44, 1-2).
Род офиоцитиум(фиг. 44, 4) има продълговати цилиндрични килийки, които могат да бъдат прави, огънати или спирално усукани, като на края могат да носят шип. Клетъчната стена се състои от две неравни части, от които повечето отучаства в растежа на клетките, като по-малката част е постоянна и има формата на шапка. Едноклетъчни и колониални видове, свободно живеещи или прикрепени към субстрата с малка дръжка. Размножават се чрез зооспори и апланоспори, срещат се цисти. Те живеят в сладки води.
Род Mischococcusобразува дървовидно прикрепени колонии. Разклоняването е дихотомно и тетрахотомно. Клетките са разположени по 2 или 4 по върховете на лигавичните разклонения на колонията. Клетките са сферични до овални, с тънка или дебела клетъчна стена. Понякога клетъчната стена е лъскава и кафява поради импрегнирането й с железни соли. Млад едноклетъчни организмис лигава, дисковидна основа, която служи като опорна точка за закрепване. След освобождаването на спорите протопластът на майчината клетка се превръща в желе и се разтяга, дължината става 6 пъти по-голяма от ширината и така се появява цилиндричен крак. Празната клетъчна стена на майчината клетка винаги става основата на стъблото. Безполово размножаване чрез зооспори и автоспори. Автоспорите са прикрепени към горния ръб на слизестата дръжка. Последващите клетъчни деления повтарят процеса и произвеждат дървоподобна колония. полов процес – изогамия. Те живеят в малки сладки водоеми като епифити на нишковидни водорасли. Известен в Централна Европа и Азия.
Разред Tribonemaceae - Tribonematales. Представителите имат нишковиден, мултифиламентозен, фалшивотъканен и тъканен тип диференциация на талуса. Клетъчните стени са или с H-образни припокриващи се части, или са твърди.
Род Трибонема- неразклонени нишки (фиг. 46, 1). Клетките са цилиндрични или бъчвовидни. Клетъчната стена се състои от две половини, които се намират с ръбове един върху друг в средата на клетката. Черупките често са наслоени. Фрагменти от нишки винаги завършват с празни половини на H-образни фрагменти от черупки, които приличат на вилица. В клетките има няколко жълтеникаво-зелени пластиди, няма пиреноиди. Размножаването е вегетативно (фрагментиране на нишките), безполово (чрез зооспори и апланоспори) и полов път (изогамия), като апланоспорите се произвеждат по-често от зооспорите. Може да образува акинет. Те живеят в сладки води, където се развиват особено изобилно през студения сезон.
Разред Vosherievye - Vaucheriales. Всички представители имат сифонален талус, оогамен полов процес и синзооспори.
Род Вошерия(фиг. 46, 3) има талус с неклетъчна структура; неговият талус достига дължина от няколко сантиметра, прикрепен към субстрата с помощта на безцветен ризоид. В нишките няма прегради, по-голямата част от талуса е заета от вакуола, а множество ядра и пластиди са разположени по периферията в цитоплазмата. Нишки с апикален растеж и рядко странично разклонение. Преградите се образуват, когато талусът е повреден и за отделяне на репродуктивните органи. Безполовото размножаване се осъществява от апланоспори, синзооспори, акинети. Синзооспорите се образуват една по една в зооспорангиума, който е отделен от вегетативните клетки чрез преграда в края на нишката. Зооспорите са многоядрени и многофлагелирани. Половият процес е оогамия. Зиготата е покрита с дебела обвивка и след период на покой прераства в нов талус.
Видове Вошеришироко разпространен в сладки, солени и морски води, както и в сухоземни местообитания. Срещат се на всички континенти, включително Антарктида. Те образуват тревисто-зелени или тъмнозелени преплетени маси - така наречените рогозки, гладки, пълзящи или възглавничести. Водни, полуводни, сухоземни форми. Те живеят в различни местообитания: морета, естуари, устия, солени блата, мангрови гори, потоци, канали, езера, езера, обработваема земя и блата.
Значениехетероконтови водорасли
От отдела за водорасли Heterocont, кафявите водорасли са от най-голямо значение за природните екосистеми и за хората.
Кафяви водорасли - осн източник на органична материяв крайбрежната зона на моретата. Тяхната биомаса в моретата на умерения и субполярния пояс може да достигне няколко десетки килограма на квадратен метър. Гъсталаците от кафяви водорасли осигуряват условия за хранене и размножаване на много крайбрежни животни и други водорасли. Чарлз Дарвин наблюдава кафяви водорасли край бреговете на Южна Америка Макроцистис, написа: „Мога само да сравня тези обширни подводни гори на южното полукълбо със сухоземните гори на тропическите региони. И все пак, ако една гора бъде унищожена в някоя страна, не мисля, че дори приблизително същият брой животински видове ще умрат, както при унищожаването на тези водорасли.
Служат гъсталаци от кафяви водорасли място за храна,подслон и отглежданемного животни. Образно казано, кафявите водорасли осигуряват на другите водни организми „маса, подслон и детска стая“.
Кафявите водорасли се използват широко от хората. Те са богати йоди други микроелементи. Народите от Югоизточна Азия традиционно ги използват за храна, особено представители на разред водорасли, от които се приготвят голямо разнообразие от ястия. Фуражно брашно, приготвен от кафяви водорасли, повишава продуктивността на добитъка; в същото време се увеличава съдържанието на йод в яйцата и млякото.
От кафяви водорасли получавамалгинати- соли на алгиновата киселина. Алгинатите се използват широко в различни индустрии. Това са нетоксични съединения с колоидни свойства, така че се използват широко в хранително-вкусовата и фармацевтичната промишленост. Алгиновата киселина и нейните соли са способни на 200-300-кратна абсорбция на вода, образувайки гелове, които се характеризират с висока киселинна устойчивост. В хранително-вкусовата промишленост те се използват главно като емулгатори, стабилизатори, желиращи и водозадържащи компоненти. Например, сух прах натриев алгинат се използва в производството на прахообразни и брикетирани разтворими продукти (кафе, чай, мляко на прах, желе и др.) За бързото им разтваряне. Водни разтвориалгинатите се използват за замразяване на месни и рибни продукти. До 30% от общия обем алгинати, произведени в света, отиват в хранително-вкусовата промишленост.
В текстилната и целулозно-хартиената промишленост алгинатите се използват за сгъстяване на бои и увеличаване на здравината на връзката им с основата. Импрегнирането на тъканите с някои алгинати им придава защитни свойства: водоустойчивост, киселинна устойчивост и повишава механичната якост. Редица соли на алгиновите киселини се използват за получаване на изкуствена коприна. По време на Втората световна война в САЩ и Англия от алгинова киселина и нейните соли се произвежда голямо количество камуфлажни тъкани и мрежи за жилищни и промишлени сгради.
Алгинатите се използват в металургията: в леярната те подобряват качеството на формовъчната пръст. Солите на алгиновата киселина се използват при производството на електроди за електрозаваряване, което позволява получаването на по-високо качество на заварките. Алгинатите се използват и в производството на пластмаси, синтетични влакна, покрития и устойчиви на атмосферни влияния строителни материали. Използват се при производството на висококачествени смазочни материали за машини. В радиоелектрониката алгинатите действат като свързващ агент при производството на висококачествени ферити.
Най-голямо приложение намира водоразтворимият натриев алгинат, способен да образува вискозни разтвори. Използва се широко за стабилизиране на различни разтвори и суспензии. Добавянето на малко количество натриев алгинат към хранителни продукти - консерви, сладолед - подобрява тяхното качество. Използва се и за направата на декоративна козметика, кремове и маски в парфюмерийната индустрия.
Във фармацевтичната индустрия алгинатите се използват за покриване на таблетки, хапчета, като компонентни основи за различни мехлеми и пасти, като гелове за носители на лекарства и при производството на разтворими хирургически конци. В медицината калциевият алгинат се използва като хемостатично средство и като сорбент, който премахва радионуклидите (например стронций). Годишното производство на алгинати в света надхвърля 20 хиляди тона.
Друго важно вещество, получено от кафяви водорасли, е шестводният алкохол манитол. Манитолът се използва като заместител на захарта при диабетици. Освен това може да се използва като плазмозаместител за консервиране на кръвта. Използва се за производство на таблетки във фармацевтичната индустрия. Манитолът се използва и при производство на синтетични смоли, бои, хартия, експлозиви, обработка на кожи.
фукоидани,получени от кафяви водорасли са ефективни антикоагуланти, дори по-активни от хепарина. Използването им за производството на противотуморни лекарства и антивирусни съединения се счита за обещаващо. В края на краищата, дори при най-ниски концентрации, фукоиданите могат да инхибират прикрепването на вирусите към повърхността на клетките. Фукоиданите също имат способността да образуват изключително силна и вискозна слуз, която се използва при приготвянето на стабилни емулсии и суспензии.
Енергийната криза, която обхвана много страни по света през последните години, доведе до необходимостта от търсене на нови нетрадиционни източници на енергия. И така, в САЩ за тази цел се проучва възможността за отглеждане на гигантски водорасли водорасли. Макроцистиспоследвано от преработката му в метан. Изчислено е, че от площ от 400 km 2, заета от това водорасло, могат да се получат 620 милиона m 3 метан.
Хетероконтните водорасли от класовете златни, жълто-зелени, синура, рафидофитни и евстигма водорасли, представени главно от микроскопични организми, са широко разпространени в сладките водоеми на всички климатични зони на земното кълбо, но са по-често срещани в умерените ширини. Сред златните водорасли има видове, които живеят в моретата и солените езера и много малко в замърсените води. Златните водорасли достигат максимално развитие през студения сезон: те доминират в планктона в началото на пролетта, късна есен и зима. По това време те играят значителна роля като производители на първична продукция и служат като храна за зоопланктонните организми.
Някои златни водорасли, например УрогленаИ Динобрион,развивайки се в масови количества, те са в състояние да предизвикат цъфтеж на водата. Те отделят алдехиди и кетони, които могат да придадат на водата неприятна миризма и вкус, а Уроглена– мастни киселини, токсични за рибите.
Рафидните водорасли са широко представени в планктона на сладководни тела с преобладаващо киселинно рН, особено в сфагнови блата, по-рядко в големи езера. В сладки водни тела могат да се образуват местни "цъфтежи". Гониостомум.Рафидните водорасли се срещат и в обезсолени морски заливи и локви на морския бряг, както и в открито море. С масово развитие в крайбрежните морски води те предизвикват токсично „цъфтене” на водата. И така, край бреговете на Канада по време на "цъфтежа" концентрацията на клетките на рафидното водорасло Heterosigma може да достигне 30 милиона на 1 литър. Избухването на рафидни водорасли често води до развитие на "червени приливи", които са свързани със смъртта на рибата. Причината за такива "червени приливи" може да бъде вид раждане Hattonella, Olistodiscus, Heterosigma и Fibrocapsa.
Водораслите Sinura с масово развитие в сладки водоеми могат да придадат на водата неприятна миризма ( Синура). Водораслите Feotamnia се срещат в застояли и бавно течащи сладки водоеми, където се установяват епифитно върху нишковидни водорасли.
Водораслите Eustigma се срещат само в прясна вода или в почвата.
Жълто-зелените водорасли са разпространени на всички континенти, живеят предимно в сладки води и почви, както и в сухоземни, соленоводни и морски местообитания. Жълто-зелените водорасли обитават чисти и замърсени води с различни стойности на pH: те могат да живеят както в кисели, така и в алкални води. Те се срещат главно в чисти сладководни резервоари, по-рядко в морета и солени води, предпочитайки умерени температури, по-често се развиват през пролетта и есента, въпреки че има видове, които се срещат през всички периоди на годината, включително зимата. Най-често те могат да бъдат намерени в клъстери от нишки и сред гъсталаци на висши водни растенияв крайбрежната зона на реки, езера, езера и резервоари.
По-голямата част от жълто-зелените са свободно живеещи форми, но има и вътреклетъчни симбионти - зооксантели в протозойни клетки. Интересна вътреклетъчна симбиоза се образува от хлоропластите на морските видове Вошерис мида Елизион. В продължение на девет месеца това мекотело е способно на фотоавтотрофно фиксиране на въглероден диоксид в културата. Това е най-дългата симбиоза от този тип, когато симбиотичният пластид е в пряк контакт с цитоплазмата на животното. В природата ларвите на мекотелите се хранят с нишки. Вошери. В резултат на фагоцитозата хлоропластите на водораслите навлизат в цитоплазмата на епителните клетки на мекотелите. По време на този процес хлоропластната мембрана става трислойна, една външна мембрана на CES (хлоропластен ендоплазмен ретикулум) се губи. Това явление е добро доказателство, че в хода на еволюцията хлоропластите с три мембрани могат да възникнат в резултат на вторична симбиогенеза поради загуба на мембрани.
Жълтозелените, златните и други хетероконтни водорасли са производители на кислород и органични вещества; те са част от хранителната верига. Хетероконтофитите участват в самопречистването на замърсени води и почви, образуването на сапропел, в процеса на натрупване на органични вещества в почвата, оказвайки влияние върху нейното плодородие. Те се използват като представителни организми при определяне на състоянието на замърсяване на водата; жълто-зелените водорасли са част от комплекс от микроорганизми, използвани за пречистване на отпадъчни води.
Контролни въпроси
Назовете характерните черти на структурата на кафявите водорасли.
Структурни особености на талиите на кафявите водорасли.
Как се размножават кафявите водорасли? Какво представляват моноспорите, тетраспорите и зооспорите, изогамията, хетерогамията и оогамията?
Какви са жизнените цикли на кафявите водорасли? Размножаване на водорасли фукус и келп.
Назовете характерните особености и типичните представители на разредите кафяви водорасли.
В какви местообитания се срещат кафявите водорасли? Какво е значението им в природата.
Стопанско значение на кафявите водорасли.
Назовете характерните черти на структурата и типичните представители на златните водорасли.
Какви пигменти и видове хранене са известни в златните водорасли?
Размножаване и екология на златните водорасли.
Назовете характеристиките и типичните представители на жълто-зелените водорасли.
Какви пигменти и видове хранене са известни в жълто-зелените водорасли?
Как се размножават жълто-зелените? Видове сексуално размножаване: изогамия, хетерогамия и оогамия?
Назовете характерните особености на структурата и типичните представители на синуричните водорасли.
Размножаване и екология на синуричните водорасли.
Назовете характерните особености на структурата и типичните представители на водораслите феотамния.
Размножаване и екология на феотамниевите водорасли.
Назовете общите характеристики на структурата и типичните представители на рафидофитните водорасли.
Размножаване и екология на рафидофитни водорасли.
Кои са общите черти в структурата и типичните представители на водораслите Eustigma.
Размножаване и екология на водораслите Eustigma.
Стойността на хетероконтните водорасли в естествените екосистеми.
МОРСКИ ВОДОРАСЛИ
основни характеристики
Водораслите са голяма група низши растенияny - от микроскопично малки, едноклетъчни организми домногоклетъчни гиганти. Терминът "водорасли" не е системаматематическа единица. Под името "водорасли" се обединяваняколко систематични разделения на нисши растения от различен произход, характеризиращи се по подобен начинживот и автотрофен тип хранене. Както подсказва името,водораслите обикновено се характеризират с воден начин на живот.
Тялото на водораслите, подобно на други по-ниски растения, еталус - талус, неразчленен на корен, стъблои лист, като при повечето видове има проста анатомична структура. Водораслите съдържат хлорофил в клетките сифотосинтезират. Структураклетки, тела, начини на размножаване са различни при отделните представителипредставители на тази група растителни отдели.
Клетките на повечето водорасли имат целулозна мембрана; има клетки, чието съдържание е заобиколено само от тънка гранична мембрана. Клетъчните стени на водораслите са разнообразни по структура и химичен състав. Основа на черупкатае протеин- въглехидратен комплекс. Черупката се характеризира с хетерогенност, наслояване. Слоевете се различават един от другдебелина, плътност и химичен състав. Често черупките са импрегнирани органични съединения(лигнин и кутин).
В черупката има специални отвори - пори. В допълнение към порите, клетъчните мембрани на много водорасли са оборудвани с различни видове израстъци.stami - четина, шипове и люспи.
В протопласта на клетката се различават ядро и цитоплазма. Правете болкаПовечето водорасли в клетката имат само едно ядро, но понякогаИма 2, 3 или повече от тях. Клетките на синьо-зелените водорасли нямат добре оформено ядро. Формата, размерът и местоположението на ядрото в клеткатаварират значително сред различните водорасли. В ядрото на водораслитеобразуват се същите структури, както в ядрата на други растения: черупка,ядрен сок, ядро, хроматинови включвания.
Цитоплазмата се състои от основното вещество (строма) и тела (органели), потопени в него. Отличителна черта на лепилототечението на водораслите е слабото развитие на ендоплазмения ретикулум.
В специфични органели - хлоропласти (наречени хромоматофори) са тела, богати на протеинови вещества,наречени пиреноиди. Пиреноидът е заобиколен от обвивка под формата на пръстен или отделни пластини, обикновено нишестениприрода. Хлоропластите съдържат зеления пигмент хлорофилсъществуващи в няколко форми.
Освен хлорофил, водораслите съдържат и други пигменти, коитокоито често маскират с присъствието си зеления цвят на водата.пораствам. Най-характерните и често срещани свиневодорасли, с изключение на хлорофил (зелен), каротин (жълт-оранжево) и ксантофил (жълто), са фикоцианин, фико-еритрин и фукоксантин. Фикоциан пигмент, разтворим във вода,оцветява водорасли в синьо (синьо-зелени водорасли);фикоеритринът, също разтворим във вода, придава червен цвят(червени водорасли); фукоксантинът причинява кафяв цвят(кафяви водорасли).
Притежавайки пигменти, водораслите дават субстрата, върху койтоте се установяват, различно оцветяване. В Антарктида например съветски учени откриха 3 езера с различен цвятвода: синьо, зелено и червено. Цветът на водата на тези езера се дължиналичието в тях на много малки водорасли със съответния цвят.Ледените брегове на Гренландия на много места с настъпването на пролеттазатоплянето му придобива червен (кървав) цвят, което се обяснява с масовото размножаване на водорасли с червен цвятсрамежлив.
Името на Червено море се дължи на растежа му в голямколичеството водорасли в това море, които имат червеникав оттенък.Различните цветове на водораслите имат адаптивна стойностnie. Дневна светлина до водорасли, потънали дълбоко във водатавинаги идва в модифициран състав. Водата е сравнително добра.предава синьо и зелено и силно абсорбира червени и жълти тоновелъчи. На голяма дълбочина, задържането на зелени лъчи с единхлорофилът е лош, червеното идва на помощпигмент, който лесно абсорбира зелените лъчи. Следователно, червеноили пурпурни, водораслите са често срещани на големи дълбочиниморета.
Водораслите са много разнообразни по форма и размер. Среща-Срещат се микроскопично малки и достигащи огромни размери (до 50 m и повече) едноклетъчни и многоклетъчни форми. Преходният етап между тях е зает от колониални водорасли.Колониите се състоят от няколко хлабаво свързани еднородниклетки. С увеличаване на размера на водораслите, някоирайска диференциация на телата им. Например в кафявите водорасли,растат големи, талусът е силно разчленен. Такива водорасли са прикрепени към субстрата с тънки безцветни нишки,които се наричат ризоиди.
Има едноклетъчни водорасли, в които тялото достигаголеми размери 0,5 м и повече. Имат голям бройядра и хроматофори. Външно тялото на такова водорасло е разчлененоно вътре няма прегради, т.е. е едноклетъчно. Пример за такова гигантско едноклетъчно водорасло ежива морска каулерпа.
Размножаването на водораслите може да бъде вегетативно, безполово и сексуално. Вегетативното размножаване се извършва на частитела на водорасли. Асексуалното размножаване възниква поради образуването на зооспори, по-рядко спори. Сексуалният процес в различни видовеводораслите е представена от: изогамия, хетерогамия, оогамия иавтогамия.
В някои по-високо организирани водорасли (червени,кафяво) има редуване на сексуални и безполови поколения.
Водораслите се характеризират с голямо разнообразие. Общброят на видовете им е над 20 хил. Класификацията на водораслите е многосложен и все още не е завършен. Видовете се обединяват в по-големи систематични единици - родове, семействаства, поръчки, класове, отдели (видове).
Цялото разнообразие от водорасли обикновено се комбинира в 6 ... 10 големи отдела (типа), представителите на които се различават по структураny, начин на размножаване, произход, но най-често предиПредставителите на тези отдели се различават един от друг по цвят.
В този учебник се разглеждат 6 отдела на водораслите: синьо-зелени, многофлагелати, диатомеи, зелени, кафяви и червениние.
Водораслите имат много древен произход, при койтомного все още не е ясно. Няма единна гледна точка за връзката между отделите на водораслите. Някои техни представителиочевидно произлизат от по-прости организми като флагелатите, а самите водорасли са предшественици на още няколко временаусукани растения (гъби, мъхове).
Представители на съвременния разнообразен свят на водораслитеимат различна древност в своя произход, те са се появили в различни геоложки епохи. Най-древните сасиньо-зелени водорасли.
Еволюцията на водораслите премина от подвижни към неподвижни форми.Разглеждат се по-примитивни и древни групи водораслитези, които прекарват живота си в мобилно състояние; повече органиНаречените водорасли се характеризират с фиксиран начин на живот. Пътекаеволюцията на водораслите идва от протозои - едноклетъчни микроскопично малки форми до многоклетъчни сложни форми.
Както вече споменахме, по-голямата част от водораслитеживеят във водна среда. Някои от тях растат в солената вода на океаните.и морета, други - в реки, езера и сладководни езера. болкавлияние върху разпространението на водораслите в дълбините на водните телаосигурява светлина, следователно, в повърхностните слоеве на водата, количествотовинаги има повече водорасли. Морски водорасличервено и кафявочесто образуват огромни подводни гъсталаци, които покриват десетки километри. В зависимост от местообитанието, водораслисе делят на 2 големи групи: бентосни и планктонни.
Бентосните или дънните водорасли живеят, като се прикрепят къмдънни или подводни обекти и образуват предимно гъсталацив крайбрежната зона. Във водата живеят планктонни водораслиокачено състояние, не са прикрепени към дъното, могат да се прехвърлятсе движат с движението на водата.
Водораслите живеят не само във вода, те се намират на повърхносттапочва, в почвата, на дървета, на купчини, но винаги влажниместа. Често, особено в началото на пролетта, почвите "цъфтят",или "позеленяват", което се обяснява с развитието на колосални колимикроскопично малки водорасли. "цъфтеж" на почвата отводораслите се срещат в различни области съветски съюз - в северната, степна и дори пустинна. Прерастване в в големи количества, създават условия за развитие на бактерии и дрмикроорганизми.
Водораслите имат голямо значениев природата и широко използванисе наричат от човек.
Като автотрофни растения, те обработват огромни коликоличеството минерали и въглероден двуокисв органична материя. Водораслите създават огромни запаси от храна за животнотосветът на моретата, океаните и сладководни резервоари. И така, 1 хектар водорасли може да даде добив, равен на 100 тона сурови или 10 тона сухимаси.
Водораслите произвеждат огромно количество кислород.
Развитието на риболова е неразривно свързано с водораслите. Като абсорбират много въглероден диоксид и отделят кислород, водораслитепречистват резервоари.
В крайбрежните страни (Англия, Франция, Норвегия, Ирландияи др.), особено в Япония, водораслите се консумират широко(морски водорасли и др.) и за фураж на добитък (суров, сух и силаженбаня). Някои водорасли се използват за торполета.
Много водорасли натрупват големи количества йод и бром. Йодът се съдържа в пепелта от водорасли около 0,2% от сухата маса.Водораслите често са основният източник на йод. Смилайки, червени и кафяви водорасли, получават ценно вещество - агар-агар,който се използва в микробиологията като хранителна средаза култивиране на микроорганизми, както и в сладкарствотопромишленост в производството на мармалад и др.
В древните геоложки епохи диатомеите заедносъс силициев диоксид образува седиментна скала - триполи, коятоизползва се в производството на динамит, тухли, за полиранепредмети и др.
В сладките водоеми водораслите участват в образуването на сапропla, или органична утайка. Сапропелът съдържа голямо количествов органични вещества и често се използва за калолечение.Сапропел, който съдържа голямо количество калциеви солийон, фосфор, желязо, могат да се използват за селскостопански фуражистопански животни.
Водораслите, особено водораслите, също могат да бъдат вредни, когатов големи количества покриват подводните части на кораба. Примасовото измиране на водораслите причинява увреждане на водата и, кактоследователно, смъртта на рибата.
Отдел синьо-зелени водорасли
Синьо-зелените водорасли или цианидите санай-примитивни и най-древни по произходорганизми. Въз основа на палеоботанически данниТъжно е, че съвременните синьо-зелени водорасли не са много по-различниот техните изкопаеми предци. В повечето случаи те са представени от едноклетъчни форми, въпреки че има и многоклетъчниnye форми, събрани в колонията.
Както показва самото име на този отдел водорасли, за тяххарактерен синьо-зелен цвят с различни нюанси в зависимост отсвързва съотношението на пигментите - хлорофил, каротин, фико-цианин и фикоеритрин.
Клетъчната мембрана на синьо-зелените водорасли се състои от пектин-vyh вещества и е покрит отвън със слизесто вещество. В клетката имkah няма морфологично различно ядро и хроматофори,цитоплазмата е импрегнирана с пигменти и затова се нарича хроматплазма.
В процеса на асимилация, вместо обичайното растително нишесте,образува се малък въглехидратен гликоген (животински нишесте).
Синьо-зелените водорасли се размножават много бързо, с простаразполовяване на клетките. Процесът им на сексуално размножаванеотсъстващ.
Според структурата и естеството на клетъчното делене на синьо-зелената вода-израстъците са подобни на бактериите. Те, подобно на бактериите, нямат ясно изразено ядро, нямат флагеларни стадии на развитие.съществуват. В някои синьо-зелени водорасли (влакнести)Намаляването става в зоните, в които тези водорасли се разпадат. Такива зони се наричат хормогония. При неблагоприятнипри приятни условия от обикновени клетки се образуват спори, коитопокрити с дебела обвивка. Защитава съдържаниетоот неблагоприятни условия, благодарение на което водораслите задържатжизнеспособност за дълго време. В началотоблагоприятни условия, спорите покълват и дават начало на новклетка.
Синьо-зелените водорасли живеят предимно в прясна вода.води - езера, езера, реки, но също и в моретата,на повърхността на почвата, на скалите. Синьозелените водорасли могатживейте сякаш ниски температурина сняг и лед и на високтемператури (до 80 °C) в горещи извори. След като умряклетки маса от водорасли под формата на люспи с мръсен зеленикав цвятизплува на повърхността на водата.
Древен произход, недиференцирана структура на лепилототечение, липса на образувано ядро и полов процес, размножаванечрез просто клетъчно делене, способността да образуват спори -всички тези характеристики сочат към примитивността на синьо-зеленотоводорасли. опростен по свой начинкъм определена структура те са значителноразлични от другите водораслиТя и най-близо до бактериите.
Синьо-зелените водорасли имат добра адаптивностустойчивост на различни условияями заобикаляща среда, кое едопринесе за тяхното опазванедо ден днешен без многопромени.
Разделяне на синьо-зелен водородsli обединява около 1400 wiдов. Представители на този отдел могат да бъдат такава водарастат като chroococcus, oscillatoria, nostoc и др.
Chroococcus- едноклетъчно водорасло Шаровидна форма, понякога тези водорасли образуват, така да се каже, колонии.Често такива водорасли се събират в групи от 2 ... 4, разделенитънка преграда и заобиколен от обща доста гъста слуздебел слой.
Chroococcus е широко разпространен в блатата сред водните раси.сенки, между неравности и кал.
Осцилаторна- нишковидни синьо-зелениводорасли, често срещани в застояли водоемиобразува тъмнозелен филм на повърхността на водата или на тинесто дъно. Клетките на това водорасло са с цилиндрична форма.свързани в една нишка.
Nostok - нишковидни синьо-зелени водорасли, нишкиили неговите вериги са свързани в колонии, често със сферична форма,с големината на плод слива. Отвън тези колонии са покрити с желатинмаса. Носток живее по бреговете на езера и езера, на влажна почва.и на дъното на водоеми.
Някои синьо-зелени водорасли, заедно с гъбичките, се образуватразлични видове лишеи.
Отдел за смесени връзки морски водорасли
Multiflagellate, или жълто-зелено,водораслите се характеризират с това, че техните зооспори имат2 неравни флагела, като късото камшиче е гладко, а дългото камшиче е пересто. Хроматофорите жълто-зелени, дисковидни.Едноклетъчни, колониални, нишковидни и неклетъчни организми.Такъв пример е ботридиумът.
Ботридиум- най-типичниятплантатор за многофлагелатни водорасли. Талусът е зелен везикул с диаметър 1-2 mm, от долната страна на везикуларазположени са безцветни разклонени израстъци - ризоиди, коитоводораслите са вградени в почвата. Това е едноклетъчно многоядреноморски водорасли. Средата на мехурчето е изпълнена с клетъчен сок, цитоплазмата е разположена париетално. Хроматофорите съдържат много каротиноиди, така че ботридият има жълтеникаво-зелен цвят.Липсват пиреноиди.
Размножава се главно чрез зооспори, които се произвеждатв колосално количество в средата на балона. Живее на влагаземята, по краищата на локвите, образува тъмнозелени набези върху почватацветове.
Отдел диатомеи морски водорасли
Диатомеи, диатомеи, или силициев диоксид, или бацили, водорасли са изключителносорт чай, микроскопично малък, най-вечеслучаи на едноклетъчни организми. Характеризира се този отдел водораслиrizovaetsya особена структура на клетките. Клетката е покрита с твърдо веществообвивка под формата на пектин, желатинообразен филм, отвънтя е покрита с кремъчна черупка, която се състои от две самостоятелнистоящи половини, така наречените крила. Едно от тезикамъкът покрива другия като капак на кутия. По половинкитечерупката има процеповидни отвори от двете страни. През негоотваряне на цитоплазмата на клетката комуникира с външната среда. мишенаki имат изключителна сила. Те не се усвояватXia животни и птици, не се унищожават дори при нажежаема жичказапален. Има над 5000 вида. Отличителни гледкиОсновните характеристики са формата на клетките и различните удебеления по тяхчерупки под формата на белези, мрежи и др.
Формата на клетките на някои видове диатомеи еудължена квадратна, елипсовидна, кръгла, звездовидна, във форматаленти, спирали и др.Клетките съдържат цитоплазма, ядро и единили повече хроматофори. В допълнение към хлорофила и фукоксантина,Хроматофорите съдържат и други жълто-кафяви пигменти,следователно хроматофорите са жълти. В клетките на диатомите няма нишесте, резервните вещества са представени отленено масло.
Размножават се основно чрез директно делене, коетовървят по свой път. По време на деленето всяка дъщерна клеткаполучава ядро, един хроматофор и само една от клапите на черупката,втората гънка се формира наново. В допълнение към директното деление, диатоморските водорасли се размножават полово, когато се слеятдве клетки, които преди това са отделили черупките си.
Диатомеите живеят в морски и сладки водичесто те са основните интегрална частпланктон и служат като ценна храна за животните. Листата на мъртвите клетки на водораслите потъват на дъното и постепенно образуват огромни отлагания,известен под името „планинско брашно, диатомит, триполи.
Големи находища на диатомеи под формата на диатомеи иla са концентрирани в полярните региони на океаните, близо до Аляска,Алеутски острови, в Охотско и Берингово море. На разположениете са в Балтийско море.
Диатомеите включват сладководните водорасли pineulusриа,фрагиларияи отчетна картария; тези водорасли образуват колонии под формата на лентиили вериги;навикуларасте храстовидно колоniei.
Отдел за зелени водорасли
Зелените водорасли са един от най-разнообразните отдели на водорасли, обединява около 5000 вида.Представителите на този отдел се характеризират със зелен цвят,което се дължи на хлорофила и не се маскира от никаквиили други пигменти. Зелените водорасли са представени от едноклетъчни, многоклетъчни, колониални форми. Зеленитеводораслите най-често имат нишковидна структура, нишките се състоят отедин ред клетки. Клетката има целулозна мембранаплазма, ядро и хроматофори. Размножаването става безполововегетативно и полововъв водата живеят зелени водорасли,Но някои от тях живеят и на сушата, натеми.
Мнозинството ще представиiley живеядървета, върху камъниnyah на влажни, сенчести места. В езера и реки формакал.
Отделът на зелените водорасли е разделен на няколко класа, от коитонека разгледаме равни флагели, или всъщност зелени, чифтосващи се водорасли или конюгати, и char, или лъчиРазред равножлезовидни Сред зелени водирастат, този клас е най-обширният. Представителите са разнообразнипо външен вид и вътрешна структура. Те са характерниналичието на две еднакви флагели Този клас съчетава единклетъчни и колониални, подвижни и неподвижни форми.Класът е разделен на 8 поръчки. Нека разгледаме представители 4 отредове.
Поръчка волвокс . Повечетотипични представители на разред Волвокс сахламидомонада и волвокс.
Chlamydomonas е едноклетъчномобилни водорасли. Живее в големи количествав сладки плитки водни тела - локви, езера, канавкипокрай пътищата. При обилно размножаване оцветява водата в зеленоцвят. Представлява микроскопично малка овална клеткаили кръгла форма. В единия (предния) край клетката е удълженапод формата на чучур, в същия край има 2 равни флагела, които допринасят за движението на водораслите. Запълва се цялата кухина на клеткатанепокрити от цитоплазма, в която се намира ядрото, по-близо до носа
Волвокс, илипумпал, е характерен пример за колониални форми на микроскопични водорасли. Колонията на това водорасло се вижда с просто око, достигас размер на глава на карфица и има формата на топка. Такава сферична колония на Volvox се състои от огромен брой клетки (до 50 хиляди), разположени в един слой по периферията на топката.Всяка клетка носи 2 флагела. Всички флагели са разположени по дължинатапериферия и улесняват движението на цялата колония. Движениекамшичетата на всички клетки винаги са координирани. Кухината на топката е запълненанепокрити с течна слуз. Волвоксът се размножава вегетативно исексуално. По време на вегетативното размножаване в рамките на майкатаколония (топка) Образуват се 8 ... 15 дъщерни колонии (топки). Приузряване на дъщерните колонии, стените на възрастното кълбо се разкъсвати излизат млади колонии, след което майчиното оцвНия умира.
Половото размножаване - оогамията - се среща само при повечетоlee по-големи клетки. В същото време в клетките на колонията Volvox, окологоли гамети с две камшичета се разделят, сливат се само гамети от различни индивиди на водораслото.
Има много видове Volvox. Всички те са широко разпространениглавно в застояла, добре загрята сладка водаводоеми със застояла вода - езера, езера, по-рядко в реки
Хлореламикроскопични едноклетъчнисферични морски водорасли. Хлорелата се характеризира с бързоразмножаване и много активен процес на фотосинтеза. Благодарение наналичието на голям брой пластиди на хлорела има различен ефектактивно използване слънчева енергия, конвенционален културенрастенията консумират само 0,1% от слънчевата енергия с биохимични трансформации и хлорела -2,5%, тоест 25 пъти повече.други положително качествохлорела - много бърз растежжени.
Ulothrix е нишковидно неразклонено водорасло,състоящ се от един ред клетки. Долна клетка с безцветен конецима особена удължена формаС тази клетка водораслото се прикрепя към подводни предмети.
Улотриксната нишка се удължава поради напречното клетъчно деленеи може да расте за неопределено време. Клетките са хомогенникъси, всяка от тях цитоплазма, ядро, хроматоформис пиреноиди. Ulothrix се размножава главно безполово.от, образувайки 4-флагелирани зооспори. Тези зооспори савремето се движи и след това се установява върху някакъв подводен обекти прерастват в нова нишка.Половият процес е изогамия. Освен това мъжките и женските гамети са външно неразличими, но физиологично те са различни и идват от различни теми, толкова различнигаметите се означават с + и -.
Ulothrix живее в прясна вода, където растеводни обекти (камъни, купчини), които стават ярко зеленицвят.
Cladophora е нишковидно зелено водорасло.Талусът се състои от разклонени нишки, образувани от един редмногоядрени клетки, отделните растения изглеждат като подводнихрасти Клетките са големи, многоядрени, с дебела,слоеста обвивка. Среща се в сладки и солени води
Дивизия червено водорасли, или алени насчиИма повече от 600 рода и около 4000 вида. от други водораслисе различават по оцветяване, което се дължи на наличиетоосвен хлорофил имат още два пигмента - фикоеритрин (червен) и фикоциан (синьо-зелен). От отношениеЦветът на водораслите зависи от образуването на тези пигменти (варира отчервено до почти черно). Представители на отдела в по-голямата част от многоклетъчнините организми сложна структура, и то само най-примитивнитеимат едноклетъчен или колониален талус. По-често талусима формата на храсти, плочи, в някои е много силно разчленен.
Червените водорасли живеят в моретата на по-големи дълбочини отзелено и кафяво. Това е така, защото червеният пигментдопринася за улавянето на големи дълбочини на зелени и сини лъчиспектър и по този начин подобрява процеса на фотосинтеза. В червеноводорасли депозит не обикновено нишесте. За разлика отобикновено нишесте, не се оцветява с йод в синьо, а в червенокафяв цвят.
Червените водорасли нямат зооспори и сперматозоиди.Безполовото размножаване при тях става с помощта на неподвижниспор. Полово размножаване - оогамия, но вместо спермаобразуват се мъжки клетки - сперматозоиди, те се пренасят в яйцата чрез вода. Ооцитите се произвеждат в специални органи -карпогони.
_ Червените водорасли включватделасерис талус под формата на храст; полисифонияпри които талусът има формата на разклонена нишка.
Отдел кафяво морски водорасли
За представители на отдел Кафяви водораслитрън кафяв цвят поради наличието в хроматофоритекафяв пигмент - фукоксантин. Наличието на фукоксантинмаскира зеления цвят и придава на тези водорасли кафяв цвятразлични нюанси. Освен фукоксантин те съдържат ксантофило и каротин. Отделът на кафявите водорасли се обединява над900 вида.
Кафявите водорасли обикновено се отличават с голям многоклетъчен талус. Най-големите представители на водораслите се срещат именно сред кафявите водорасли. Някои от тях, като макrocystis, достигат до 60 м дължина, но има и малки формина няколко милиметра една от друга.
Вместо нишесте клетките на кафявите водорасли съдържат глюкозаза и захарни вещества - примамва и водорасли, които даватТези водорасли имат сладък вкус при готвене. Каторезервно вещество те често отлагат масла.
Талусът на кафявите водорасли е многогодишен, но листенплочите умират ежегодно и растат отново в началото на пролетта.
Сложната външна структура на кафявите водорасли определяпритях и диференциация в анатомичната структура (иматразлични клетъчни форми). Някои учени смятат, че тезиводораслите дори имат различни тъкани.
Кафявите водорасли се размножават по различни начини. Неконякои от тях се размножават по примитивен полов път – изmia, когато 2 гамети с еднаква форма се сливат. други,по-развити водорасли (келп) има по-сложенполов процес - оогамия, при която голямо яйцесе слива с малка подвижна мъжка гамета - сперматозоидикъща.
При кафявите водорасли се случва безполово размножаванезооспори, които се произвеждат в големи количества в зооспоредици. При кафявите водорасли редуването на поколенията е доста ясно изразено; асексуални и сексуални. На листни плочитези водорасли образуват едноклетъчни зооспорангии, събраниnye на групи, сред които са безплодните нишки.Всеки зооспорангий произвежда 16...64 или повечезооспори. Зооспорите са външно идентични, но физиологично различни.Някои от тях покълват и образуват микроскопично малки съпруги.скай, а други - мъжки гаметофити. върху мъжките гаметофитидопълнително се образуват антеридии и в тях един сперматозоидзоид, а върху женските гаметофити се образуват оогонии, носещиедно яйце. След сливането на спермата и яйцеклеткатаобразува се зигота, от която се развива безполовото поколение -спорофит.
Кафявите водорасли са морски обитатели, много отте са по-често срещани в северните морета, често се образуватогромни гъсталаци в моретата и океаните. Северно от Атлантидаокеан, в Саргасово море, в огромен бройИма род кафяви водорасли - Sargassum. Тези водорасли са повечеот всички са в плаващо състояние поради наличието наспециални мехурчета, пълни с въздух.
Кафявите водорасли се считат за древна група растения, те иматустановява се по-висока степен на диференциация не самовъншната, но и вътрешната част на талуса. Външно те са подобни наshimi растения, така че някои ботаници смятат, че тезиводораслите може да са дали начало на висши растения.
Отделът на кафявите водорасли се състои от 4 разреда. Помислете предипутери от два реда: водорасли и фукус.
Поръчайте водорасли. Товамного големи водорасли, понякога достигащи 60 m или повече.Техният талус е силно разчленен и освен това е добре развитразклонени ризоиди, с които водораслите са здраво прикрепени към дъното на морето. Ламинарията живее в крайбрежната ивица на моретатана дълбочина 5 ... 10 m и често образуват огромни подводни гъсталаци"гори".
Ламинарите включват род Laminaria (включва 30 вида), род Lessonia (включва5 видове) и рода Macrocystis. Тези водорасли са многогодишни растения, които се различават едно от другоструктура на талуса.
Отдел слузести плесени
Плазмодият се образува от сливането на голи амебиразлични клетки на слузестата плесен и при някои видове слузести плесени достигаразмер на човешка длан.Той обикновено оцветени в ярко жълтоцвят и има способността да много бавно прилича на амебадвижение (0,1 mm/min). Когато се движи, плазмодият има тенденция даскрийте се от светлината и се насочете към източника на влага. Срещаслузните плесени обикновено се срещат в сенчести гори; на гнили растения, междукора и дърво, в напукани пънове, под паднали листа.Размножават се чрез спори.
Позицията на слузестите плесени във филогенетичната система е неясна,очевидно те са произлезли от някакъв вид флагела. От товаотдел, разгледайте плазмодиофора.
Болестта се разпространява през почвата и се развива особено бързо.вирее на кисели почви.
Всички хетероконти изглежда имат флагеларен апарат. Камшичетата са 2, като на единия от тях има много характерни тръбести тричленни перести израстъци, или власинки - мастигонеми. Наличието на две неравни флагели послужи като основа за наименуването на тази огромна група водни организми многофлагелативодорасли, или хетероконтами. В допълнение към разликите в дължината, камшичетата се различават и морфологично: основният флагел се състои от ос и ресничести косми, разположени на перисти върху него, страничният флагел е гладък и флагелат. Съставът на пигментите също е подобен - хлорофили АИ си множество каротеноиди, както и структурата на хлоропластите. Хлоропластите са заобиколени от четири мембрани, като двете външни мембрани са свързани с ендоплазмения ретикулум на клетката.
Като част от отдела за водорасли Heterocont (Diversified), няколко групи водорасли са обединени в ранг на класове: кафяви, златни, жълто-зелени, Rafidophytes, Sinura, Feotamnia, Eustigma, някои от които преди са били считани за независими отдели.
Клас Кафяви водорасли– Phaeophyceae (Fucophyceae)
Името на класа "кафяви водорасли" отразява цвета на техния талус (от гръцки. Рhaios- мургав). Цветът се дължи на наличието на голям брой кафяви и жълти пигменти - в допълнение към хлорофилите АИ сИ β -каротин в техните хлоропласти има излишък от ксантофили, особено фукоксантин.
Многоклетъчни, предимно големи водорасли, особено широко разпространени в студените води на двете полукълба. Сред кафявите водорасли не са известни нито едноклетъчни, нито колониални форми. Известни са около 1,8 хиляди вида.
Клетъчна структура. Клетката при видовете кафяви водорасли е еукариотна: има образувани органели със собствени мембрани: ядро, митохондрии, пластиди и др. Зелени пигменти - хлорофили АИ с- маскирани от допълнителни: оранжеви каротини и жълто-кафяви ксантофили; от последните преобладава фукоксантинът. В клетките на кафявите водорасли се откриват един до много пластиди. Върху външната клетъчна мембрана клетките на кафявите водорасли обикновено развиват твърда клетъчна стена.
Ядрото най-често е едно, но понякога има няколко ядра. При някои представители с увеличаване на възрастта на клетката броят на ядрата се увеличава. В ядрото винаги има едно ядро.
Хлоропластите на вегетативните клетки на кафявите водорасли имат око (стигма) и пиреноиди, които могат да бъдат един или повече; някои видове нямат пиреноиди. Оцелусът, състоящ се от 30-40 липидни глобули, функционира като леща, която фокусира светлината върху флагеларното издуване, което е фоторецептор. Зооспорите и гаметите на кафявите водорасли обикновено нямат око. Резервните вещества се отлагат в пиреноидите. Хлоропластите най-често са малки, дисковидни, париетални. Тяхната форма може да бъде звездовидна, лентовидна или ламеларна. Формата на хлоропластите в клетките на кафявите водорасли може да се промени с остаряването на клетката. Обвивката на хлоропласта се състои от четири мембрани, като двете външни мембрани са свързани с EPS, ендоплазмения ретикулум на клетката.
Целулозата, която изгражда клетъчната стена на кафявите водорасли - понякога наричана алгулоза - се различава по свойства от целулозата на висшите растения. Извън целулозната стена има силно слузест пектинов слой, образуван главно от алгинова киселина и нейните соли и съединения с протеинови вещества. Алгинова киселинасъщо се среща само в кафявите водорасли.
Основни характеристики.
Всички кафяви водорасли са фототрофи. Продукти на фотосинтезата - специфичен въглехидрат хризоламинарин,алкохол манитолИ липиди.Липидите се натрупват под формата на маслени капчици. Хризоламинаринът се отлага извън хлоропласта: той се намира в специални вакуоли в цитоплазмата близо до пиреноида. Манитолът изпълнява резервна, както и осморегулаторна функция; концентрацията му в клетката зависи от промените в солеността на водата.
Младите клетки на кафявите водорасли съдържат малки и многобройни вакуоли, които стават по-големи с възрастта поради сливането. В цитоплазмата има специални образувания – физоди, които съдържат флоротанини.Тези вещества са известни само при видовете кафяви водорасли. Функцията на флоротанините може да бъде свързана със защитата на талиите на водораслите от изяждане от животни, като мекотели. Флоротанините на кафявите водорасли вероятно инхибират заселването на епифитни водорасли и животни на повърхността на талуса. Предполага се, че флоротанините участват в защитата срещу радиационни увреждания и в селективната абсорбция на йони на тежки метали от водната среда. Безцветните флоротанини се окисляват във въздуха, за да образуват пигмента фикофеин, който придава характерния тъмен цвят на изсушените кафяви водорасли.
Структурата на талуса.Сред видовете кафяви водорасли няма едноклетъчни и колониални форми, всички индивиди са многоклетъчни. Формата на талиите на кафявите водорасли е най-разнообразна. Това са нишки, пълзящи или вертикално стоящи, едноредови или многоредови; масивни твърди или керемидени корички и корички; тънки лигавични плочи или плътни кожени плочи на стъблото, прости или разчленени; мехурчета и торбички; тръби и въжета; различно разклонени тънки храсти или мощни храсти. Закрепването на талиите се извършва с помощта на ризоиди или подметки. За да ги поддържат във вертикално положение, големите бентосни форми на кафяви водорасли образуват въздушни (плуващи) мехурчета, пълни с газ (фиг. 29).
Ориз. 29. Външен вид на кафяви водорасли: 1 - Fucus; 2 - Захарин; 3 - Алария; 4 - Саргасум
Таломите на представителите на разредите Laminaria и Fucus са най-сложните сред видовете кафяви водорасли. Те имат признаци на тъканна диференциация със специализация на клетките на талуса (фиг. 30). В техните тали се различават корасъстоящ се от интензивно оцветени клетки и сърцевина, състоящ се от безцветни клетки, които често са събрани във влакна. При Laminaria в сърцевината се образуват и ситовидни тръбички и тръбести нишки. Ядрото изпълнява не само транспортна функция, но и механична, тъй като съдържа нишки с дебели надлъжни стени. Между кората и сърцевината на много кафяви водорасли може да има междинен слой от големи безцветни клетки.
Височинаталусът при представителите на кафявите водорасли е най-често интеркаларен (интеркаларен) и апикален (апикална), рядко базална. Интеркаларният растеж може да бъде дифузен или може да има зона на растеж. При големите форми интеркаларната (интеркаларна) меристема или активно деляща се група клетки се намира в точката на прехода на "петурата" в "листното" острие. Големите водорасли също имат меристематична зона на повърхността на талуса, така наречената меристодерма (вид аналог на камбия на висшите растения).
размножаванев кафявите водорасли, вегетативни, безполови и сексуални. Гаметите се образуват в многоклетъчни гаметангии, зооспорите - в 
Ориз. 30. Вътрешно устройство Ламинария(по: L.L. Velikanov et al., 1991): А- надлъжен разрез на стъблото; б- напречен разрез на стъблото; IN- срез от плоча със соруси на зооспорангии
едноместни спорангии.
Вегетативното размножаване при редица видове кафяви водорасли може да се извърши от местата на талуса; във видовете мил Сфацелария- плодови клонки (плодови пъпки); при видове от рода Фукус- група клетки на подметката, които са способни да се диференцират в нов талус.
Безполовото размножаване на кафявите водорасли обикновено става с помощта на зооспори. Някои видове се размножават чрез неподвижни тетраспори или моноспори. Спорите на асексуалното размножаване се образуват в резултат на мейоза и последващи митози в едноместни спорангии. Половият процес е изогамия, хетерогамия и оогамия. Гаметите се образуват в полиедъртези гаметангии- специални групи клетки. Във всяко „гнездо“ на гаметангиума се образува по една гамета.
За кафяви водорасли, полови феромони- разтворими вещества, които координират дейността на клетките по време на възпроизводството. Феромоните, от които са известни поне десет, могат или да стимулират отварянето на антеридии, или да привлекат мъжки гамети към женски. Те също участват в изолацията на видовете. Смята се, че феромоните се образуват в повечето видове кафяви водорасли, които имат полов процес.
Жизнен цикъл.Кафявите водорасли имат два основни вида жизнен цикъл. Един вид е хаплодиплобионт. Спорофитите са диплоидни, т.е. Всички клетки съдържат пълен набор от хромозоми. Спорите на асексуалното размножаване се образуват в едноместни спорангии. По време на образуването на спорите настъпва редукционно делене - мейоза. Такова намаление се нарича спорно. Образуваните след мейоза клетки (спори) са хаплоидни; съдържа половината от броя на хромозомите. Хаплоидните зооспори и тетраспори покълват в хаплоиден гаметофит, върху който се образуват гамети в многоклетъчни гаметангии. Диплоидната зигота, образувана след сливането на гамети, прераства в диплоиден спорофит.
Друг тип е диплобионт; спорофитите, спорите и гаметофитите са диплоидни; редукционно делене възниква по време на образуването на гамети. Такова намаление се нарича гаметично. След сливането на хаплоидни гамети се образува диплоидна зигота, която покълва в диплоиден спорофит. Този тип е често срещан при видовете водорасли (фиг. 31).

Ориз. 31. Жизнен цикъл Захарини(по: L.L. Velikanov et al., 1991): 1 – спорофит, 2 – соруси на зооспорангии, 3 – зооспори , 4 - гаметофит, 5 - антеридий, 6 - оогониум, 7 - антерозоид, 8 - зигота (оплодено яйце)
На макроскопичен спорофит Захарини, в горната част на плочата се развиват ивици сори(групи) от зооспорангии. В зооспорангиите след редукционно делене се образуват зооспори с две неравни камшичета. Те растат в микроскопични гаметофити: мъжки с антеридии и женски с оогонии. Яйцето, оплодено от антерозоиди, веднага, без период на сън, покълва и образува спорофит. В този жизнен цикъл Захаринизавършва.
Водораслите Fucus нямат спорофити. При Фукусамъжките и женските полови органи - антеридии със сперматозоиди и концептакули с оогонии и яйца - се намират във вдлъбнатините на талуса - скафиди, мъжките и женските скафидии могат да бъдат на едно и също съдили в различни рецепти. Oogonia седи на клетка-крак , появяващи се на стената концептула(фиг. 32, C, D).Оогониевата клетка претърпява три деления

Ориз. 32. Размножаване Фукуса(според: B. Fott, 1971; W. Braune et al., 1974): А – фрагмент от талус (1 – въздушно мехурче, 2 - рецепта); б– разрез през съдовете; IN– разрез на женска скафидия; Ж- оогония; д- освобождаване на яйца; д– разрез на мъжки скафид; И– клон с антеридии; 3 – продукция на сперма
ядра, като първото от тях е редукция, в резултат на което в оогония се образуват 8 яйца.
Антеридиите обикновено се образуват върху разклонени нишки, които ги поддържат - парафизи(фиг. 32, Д,И).В резултат на няколко деления се образуват 64 сперматозоида. сперматозоиди Фукусаимат око, разположено вътре в редуциран хлоропласт. В предния край на сперматозоида се образуват 13 микротубула скитниктекущ- специална структура, която трябва да участва в дефинирането на женския полов атрактант.
Сперматозоидите се привличат към яйцето Фукуса fucoserraten, неспецифичен феромон, секретиран от яйцето. Разпознаването на видово ниво е свързано със специфични олигозахариди в яйцето и спермата. На повърхността на яйцето има различни области, всяка от които съдържа специални органични съединения - гликопротеини. Сперматозоидите съдържат гликопротеини, организирани в места върху мембраната и израстъците на предния камшик, както и на повърхността на самия сперматозоид. При среща с яйцеклетката сперматозоидите се движат по черупката на яйцеклетката, сякаш я изследват с помощта на предния си флагел. Ако гликопротеините на сперматозоида са комплементарни на гликопротеините на яйцеклетката, тогава настъпва само разпознаване на сперматозоида от яйцеклетката.
Систематика
Фосилни находки, които може да са свързани с кафяви водорасли, са известни от средния силур (около 425 Ma). Но тези находки също са подобни на някои съвременни зелени и червени водорасли. Фосилни находки, които определено могат да бъдат свързани със съвременните кафяви водорасли, датират от миоцена (5–25 милиона години). Тези водорасли напомняща на съвременната Dictiota. Възрастта на кафявите водорасли, определена чрез молекулярни методи, е 155–200 милиона години.
Типът организация на талуса, наличието или отсъствието на пиреноид, начинът на растеж, типът на сексуалното размножаване (изогамия, хетерогамия, оогамия) и жизненият цикъл се използват за разграничаване на разредите на кафявите водорасли. През последните години системата на кафявите водорасли беше активно преразгледана. В различни класификационни схеми, като част от клас Кафяви водорасли, се разграничават от 7 до 14 разреда.
Десмарестовски орден–Desmarestiales. Водораслите са храстовидни, многоразклонени, с щипка, плосък или сплескан талус. Захваща се за основата с добре развита подметка. На повърхността на талуса се развиват косми.
Род Десмарестия (фиг. 33). Кафяви или маслинови храсти с добре очертана главна издънка, до 0,8–1 m височина. Прикрепени с конусовидна подметка. Разклонение 3-4 реда, редуващи се или срещуположни. Клоните на последните разреди са плоски или по-рядко нишковидни. Видовете са морски, живеят в арктическите и умерените ширини на дълбочина 1–20 m, образуват гъсталаци или растат поединично.

Ориз. 33. Десмарестия (р. Десмарестия)
Ралфсов орден–Ralfsiales. Представителите на разреда имат талус под формата на кори с различна структура или се състоят от основна кора и вертикални издънки.
Род Аналипус.Талусът се състои от многогодишна коровидна основна част и от нея излизат едногодишни маслиненокафяви вертикални издънки с височина до 40 см. Многоклетъчните спорангии са с цилиндрична форма и двуредово устройство. Едноместните спорангии са овални и се развиват сред асимилационните разклонения. Видовете от рода са широко разпространени в моретата на умерените ширини, образувайки пояси в приливната зона.
Поръчайте Dictyota–Dictyotales.За видовете от разреда е характерен апикален растеж. Вегетативните клетки съдържат множество дисковидни хлоропласти без пиреноиди. Характерно е паренхимен талус, изоморфно редуване на поколенията. Половият процес е оогамен, безполово размножаване чрез неподвижни спори, сперматозоидите имат само един флагел. Камшикът е дълъг, с перести израстъци и насочен напред. Представителите на разреда се срещат предимно в тропическите и по-рядко в умерените морета.
Род Dictyota (Приложение, 4A). Има талус под формата на плоски маслиненокафяви дихотомично разклонени храсти с височина до 11–15 см. Клоните са линейни, тесни, без жилки, с разклонени върхове. Тетраспорангиите образуват соруси на повърхността на талуса, които имат неопределена или пръстеновидна форма. Разпространен в морските води на умерени и топли ширини.
Поръчайте Ламинария–Laminariales. Ламинарията има талус, състоящ се от плоча, стъбло и прикрепващи органи - подметка или ризоиди. Плочата е равномерна или набръчкана, цяла или разсечена, с дупки, гънки, шишки или без тях, дълга от няколко десетки сантиметра до 20–40 м. Стъблото е разклонено или по-често неразклонено. Спорофитите са едногодишни или многогодишни, някои видове са на възраст 11–18 години. Слизестите канали са разположени в кората на много водорасли.
Най-популярните представители на кафявите водорасли са келп (видове род Ламинария или захарина), или "морски водорасли", които са разпространени предимно в Тихия океан. Те се намират от повърхността (ръба) на водата до дълбочина 200 м. Ламинарията расте за предпочитане на дълбочина 2–20 (30) м, на места с постоянно движение на водата, прикрепени към камъни и скали. Ламинарията образува водораслови пояси по крайбрежието на моретата на Далечния изток. Най-мощните гъсталаци се намират близо до Курилските острови и на около. Хокайдо, където са концентрирани около половината от световните ресурси на ламинария. Годишната световна реколта от водорасли водорасли достига 2 милиона тона прясно тегло, докато повече от 1 милион тона идват от производството на марикултура в Китай.
Род Захарин (ламинария).Талусът е дълъг от 0,5 до 5–10 m, има дебела лъскава пластина, еластично стъбло и плътно преплетени ризоиди, които са прикрепени към различни твърди субстрати. Ръбовете на плочата са гладки или вълнообразни, често има издутини и гънки по повърхността на плочата. От двете страни на плочата се образуват натрупвания на спорангии - соруси, след освобождаването на спорите горната част на плочата се унищожава. Живее от две до няколко години. Захарините са ценна промишлена суровина и обект на марикултурата. захарина японскаобразува гъсталаци в северната половина на Японско море, както и близо до южното и югоизточното крайбрежие на Сахалин и Южните Курилски острови. захарина японскае най-ценният търговски вид, има отлични вкусови качества и се използва за приготвяне на различни ястия, консерви, сладкарски изделия.
Разред Sphacelariae–Sphacelariales.
Кафявите водорасли, принадлежащи към този разред, имат разклонен талус с апикален растеж. Космати клони. Вегетативните клетки имат множество хлоропласти без пиреноиди. Жизненият цикъл на мнозинството с изоморфна смяна на поколенията, сексуалният процес е изогамия или хетерогамия. Водораслите образуват малки кафяви гроздове до 3 см височина и разклонени храсти до 20 см височина върху камъни или други водорасли; има ендофитни представители.
Род Сфацелария. То има тънък талус. Разклонените нишки образуват малки, до 0,5–3 cm високи, кафяви снопчета. Вегетативното размножаване се извършва от специализирани, често трилъчеви разклонения - пропагули. Половият процес е изогамен. Женските гамети секретират феромоните ектокарпен и мултифиден. Видовете от рода са разпространени в моретата както на полярните, така и на тропическите ширини.
Разред Сцитосифон–Scytosiphonales. Представители с чревен, мехурчест, пластинчат или торбовиден, често кух талус с дължина до 25 cm. Кората е едноредова или двуредова, вътрешната част (стена) на талуса е образувана от няколко слоя големи клетки. На повърхността на талуса се развиват многоклетъчни спорангии, които са събрани в соруси, които поддържат техните нишки - парафизии снопчета косми.
Род Сцитосифоне тънкостенна кафява тръба с дължина 4–25 cm (Приложение 4B). Талусът е прикрепен с малка подметка и има редки напречни стеснения. Среща се в крайбрежната зона на моретата на различни географски ширини, често образува пояси в приливната зона.
Фукус ред–Фукалес. Имат многогодишен талус под формата на големи неравномерно или дихотомно разклонени храсти с дължина до 1–3 m. Те са прикрепени към субстрата с конусовидна подметка. Талусите имат издънки, филоиди (листовидни образувания), вместилища (вместилища на гамети) и плувни (въздушни) мехури. Те се срещат в приливната зона на всички умерени и арктически морета, където образуват мощни гъсталаци - пояси.
Род саргасъм(фиг. 34, IN). Обилно разклонени маслиненокафяви или кафяви храсти с височина 0,5–1,5 м. Прикрепени с конична подметка. Филоидите се развиват върху странични издънки, излизащи от цилиндричното стъбло в долната и средната част на талуса. Филоидите са кожести, с централно ребро, с дължина 0,2–2 cm. Плувните мехури са овални или заоблени, със или без израстъци. Sargassums са широко разпространени в умерените и субтропичните ширини от долното крайбрежие до дълбочина 10 m, където образуват гъсти или редки гъсталаци. В южната част на Тихия океан - така нареченото Саргасово море - те образуват обширни плаващи натрупвания, в които се размножават само вегетативно.
Род Фукус (фиг. 32).Талус кожен, храстовиден, разклонен дихотомно, висок 25–50 см. В близост до брега на далекоизточните морета расте няколко години, живее в крайбрежната зона. Често образува големи гъсталаци в плитки води. Видовете от този род се характеризират с дихотомия

Ориз. 34. Появата на водорасли фукус: А – Аскофилум; б – Cystoseira; IN – саргасъм; 1 - съд, 2 - плувен мехур
разклонен талус с плоски клони с надлъжно ребро, който е прикрепен към камъните с конична подметка. Фукусите се използват като торове, като храна за добитък, за производство на фуражно брашно, алгинати, използвани в медицината за лечение на гуша, корекция на теглото и др.
Разред Ectocarps–Ectocarpales. Водораслите, които са включени тук, са микроскопични, свободно живеещи, епифити, епизоити или ендофити. Репродуктивните органи са представени от едноместни (спорангии) и многоместни (гаметангии) спорангии; биват дългоцилиндрични, късоцилиндрични или шушулковидни.
Род ектокарпус. Талус под формата на меки кафяви храсти до 2 см височина. Храстите са оформени от едноредово, последователно, дихотомно или неправилно разклонени нишки. Многоклетъчните спорангии са шушуловидни, образувани отстрани на страничните разклонения на 1–3-клетъчна дръжка. Видовете са разпространени в моретата на различни географски ширини, в чисти и замърсени води, в литорала и в горната част на сублиторала. Епифитите се срещат и в обрастването на антропогенни субстрати.
Екология
По-голямата част от кафявите водорасли живеят в морските води. В сладките води се срещат само 8 вида. Има едногодишни и многогодишни видове, чиято възраст може да достигне 15-18 години. Ламинария водорасли НереоцистисИ Макроцистис, които живеят по тихоокеанското крайбрежие на Северна Америка, имат максималния размер за всички водорасли - до 30–40 m дължина. Те образуват гигантски подводни гори в моретата на Далечния изток. Кафявите водорасли се прикрепят към голямо разнообразие от субстрати - скали, камъни, чакъл, раковини и раковини на морски животни и други водорасли. Някои малки форми на кафяви водорасли живеят в тъканите на други водорасли като ендофити. Повечето кафяви видове живеят в прикрепено състояние. Откъснатият от земята талус се отнася от течението до спокойни места с тинесто дъно и продължава да съществува там. Видовете с въздушни мехурчета върху талуса, откъсвайки се от земята, изплуват на повърхността, образувайки големи плаващи клъстери, особено в райони със стабилно кръгово течение, като в Саргасово море. Те съществуват в такива клъстери доста дълго време и се размножават само вегетативно.
Кафявите водорасли са широко разпространени във всички морета, но най-голямо развитие достигат в моретата на умерените и субполярните ширини. Техните големи тали растат прикрепени към скали и камъни, а на тихи места близо до брега и на големи дълбочини те дори могат да се държат върху клапите на черупките на мекотели и чакъла. Можете да ги срещнете на различни дълбочини - от крайбрежната зона, където при отлив те са извън водата с часове, до 40–200 м. В района на Хавайските острови, на дълбочина 180 м, видове от рода саргасъм, А Келп V Адриатическо море е открито на дълбочина 200 м. Най-изобилните гъсталаци от кафяви водорасли се наблюдават на дълбочина 6–15 м, където най-добрите условия на осветление и постоянното движение на водата, което носи хранителни вещества на техните тали и граници заселване на тревопасни животни - фитофаги.
Клас Златни водорасли -Chrysophyceae
Едноклетъчни, колониални или многоклетъчни организми, обикновено имащи флагеларен стадий в своя жизнен цикъл. Клетките са едноядрени, с един или повече хлоропласти. Обикновено има стигма (фиг. 36). Камшичетата 1–2, неравни. Съкратителната вакуола е разположена в предната част на клетката. Външните обвивки на клетките на тези организми са много разнообразни. При най-примитивните видове клетката е облечена в деликатен перипласт, който позволява образуването на издатини с различна форма (ризоподии, псевдоподии), при други плазмалемата е покрита с твърда целулозна мембрана, понякога силно слузеста. Къщите в златните водорасли се предлагат в различни форми: във формата на ваза, сферична, яйцевидна, цилиндрична; с една или повече дупки. Клетката е прикрепена към основата на къщата с гъвкав крак или може да бъде свободна. При някои хризофити върху клетъчната повърхност се отлагат варовити образувания - коколити - с различни форми и размери, а силициевите флагелати имат вътрешен силициев скелет. Разнообразието на структурата на тези скелети дава възможност да се използват останките от различни форми на изкопаеми силикофлагелати за определяне на възрастта на геоложките скали, които ги съдържат. При по-силно развитите представители на отдела на златните водорасли клетките са облечени в черупка, състояща се от силициеви люспи (понякога носещи шипове) или затворени в къщи, през отворите на които излизат флагели или псевдоподии.
Къщите при видовете златни водорасли са във формата на ваза, сферични, яйцевидни, цилиндрични, с една или повече дупки. Клетката е прикрепена към основата на къщата с гъвкав крак или може да бъде свободна.
Пигменти - хлорофили АИ с, фукоксантин. Хризофитите се характеризират с най-високо съдържание на фукоксантин сред хетероконтите (55-93% от всички каротеноиди).
Повечето видове са сладководни фототрофи, но се срещат и хетеротрофи и фаготрофи.
Резервни вещества - хризоламинарин и масла.
Сред едноклетъчните форми на златните водорасли преобладават сладководни планктонни видове, а сред многоклетъчните форми - бентосни или епифитни. Срещат се главно в хладни морски и сладки водоеми. Известни са около 360 вида.

Ориз. 36. Златни водорасли (по: C. Hoek van den et al., 1995): А- клетъчна структура; б- коренова система; IN- трипартитен мастигонема; Ж- разрез на част от хлоропласта; 1 - базално тяло; 2 - базално подуване; 3 - къс флагел; 4 - стигма; 5 - ризопласт; 6 - ядро; 7 - ядрена мембрана; 8 - ядро; 9 - митохондрия; 10 - вакуоли; 11 - слуз; 12 - плазмалема; 13 - везикули; 14 - лигавично тяло; 15 - липиди; 16 - хлоропласт; 17 - апарат на Голджи; 18 - контрактилна вакуола; 19 - дълъг камшик: 20 - 1-ви корен; 21 - 2-ри гръбнак; 22 - 3-ти гръбнак; 23 - 4-ти корен; 24 - хлоропластен нуклеоид; 25 - HES; 26 - хлоропластна обвивка; 27 - поясна ламела; 28 - ламела; 29 - дълга странична нишка; 30 - къса странична нишка; 31 - крайна нишка; 32 - тръбна част на мастигонема
структура на талусав златните водорасли най-често е монадичен, но може да бъде много разнообразен: кокоиден, палмелоиден, плазмоиден, амебоиден, нишковиден, паренхим (фиг. 37).

Ориз. 37. Златни водорасли (по: L.L. Velikanov et al., 1981): А – Охромонас: 1 - външен вид, 2 - киста; б – Хромулина: 1 - външен вид, 2 - киста, 3 - схема на образуване на филм от кисти; IN – Динобрион: 1 - общ изглед на колонията, 2 - киста; Ж – Хризамеба; д – гибердия; д – Хидрурус: 1 - общ изглед на колонията, 2 - връх на клона, 3 - зооспора
размножаване. При златните водорасли има три начина на размножаване: вегетативен, асексуален и сексуален.
Вегетативно размножаваневъзниква чрез надлъжно клетъчно делене или разпадане на колония на части поради клетъчно делене в една, две или три посоки.
полов процес(клетките на златните водорасли са диплоидни) - изогамия или конюгация. В резултат на половия процес, както и при неблагоприятни условия се образуват кисти с плътна обвивка, съдържаща силиций.
безполово размножаванеизвършва се от едно- или двуфлагелирани зооспори, които се развиват в обикновени вегетативни клетки или зооспорангии. Сексуален процес: холо-, изо- и автогамия. Хологомията е вид полов процес, при който две клетки, подобни на вегетативните клетки, се сливат в предните краища в двуядрена зигота, която след това се превръща в силикатизирана киста.
Систематика
От златните водорасли в края на миналия век въз основа на резултатите от молекулярно-генетични изследвания е изолиран клас Sinuraceae. Разграничава се и клас Feotamniaceae - от жълтозелени и златни водорасли. Поради това обхватът на клас Chrysophyceae е намален и в момента включва около 360 вида, разпределени в три разреда: Chromulinales, Hibberdiales и Hydrurales. Формата, структурата и броят на камшичетата, както и пигментният състав и структурата на тялото са важни систематични характеристики.
Разред хромулин - Chromulinales. Този ред включва организми с монаден, палмелоиден и амебоиден тип диференциация на талуса. Монадични клетки с един флагел, видим под светлинен микроскоп.
Род Хризамеба(фиг. 37, А) - сладководни амебоидни водорасли, те имат флагеларен стадий в жизнения си цикъл. Зиготата се развива в стоматоциста.
Род Хромулина(фиг. 37, IN) - едноклетъчни свободно плуващи организми, които живеят предимно в сладки води. Клетките са голи; има един или два златни пластида.
Род Динобрион(Фиг. 37 , G)- едноклетъчни и колониални, свободно плаващи или прикрепени представители. Монадите са разположени в къщи с форма на ваза, образувани от целулозни микрофибрили. В допълнение към целулозата, къщата съдържа и голям брой аминокиселини. Храстовата форма на колониите е свързана с метода на вегетативно клетъчно делене, когато една от дъщерните клетки, напускайки родителската къща, се прикрепя към дупката си. Въртейки се около оста си, тя образува собствена къща. И двете дъщерни клетки могат също да напуснат родителската къща. Клетките съдържат 1-2 хлоропласта; в предния край на клетката се виждат око и 2 контрактилни вакуоли. Хризоламинаринът се намира във вакуола в задния край на клетката. Размножаването е вегетативно и полово.
Разред Gibberdiae–Hibberdiales. Представителите на този ред се характеризират с уникален пигментен състав. В допълнение към фукоксантина, те имат допълнителен светлосъбиращ каротеноиден пигмент, антераксантин.
Включва раждане Gibberdia, Chromophyton, Styloceras, Chrysopixis, Platytekaи други.
Род гибердияима два етапа в жизнения цикъл: колониален палмелоиден неподвижен и едноклетъчен монаден подвижен. При светлинна микроскопия се вижда само един флагел в монадния стадий. В допълнение към фукоксантина, характерно е наличието на друг допълнителен светлосъбиращ каротеноиден пигмент, антераксантин.
Поръчайте Hydrorus–Hydrarales. Представителите на този ред имат талус с палмелоиден и псевдопаренхимен тип структура. Характерно е наличието на уникални тетраедрични зооспори. Няма око. Митозата е полузатворена.
Включва раждане Hydrorus, Celloniella, Federmatium, Chrysonebulaи други.
Род Хидрурус(фиг. 37, д) е най-диференцираният сред палмелоидните водорасли. Талусът му представлява големи, до 30 см дълги лигави колонии, които приличат на кафяви разклонени нишки, често излъчващи много неприятна миризма. В колониите може да се направи разлика между главния ствол и страничните клони. Клетките, потопени в обикновена слуз, съдържат един чашковиден хроматофор и няколко контрактилни вакуоли. Клетките са рехаво разположени по периферията на колонията и по-плътни в средата. Талусът може да расте само чрез делене на апикални клетки. При безполово размножаване в клетките на страничните разклонения на колонията се образуват зооспори. Образува сферични кисти. Среща се в планински потоци и реки със студена вода, където се прикрепя към твърди субстрати.
Клас Синура водорасли -Sinurophyceae
син- заедно и ура- опашка. Тази група организми е изолирана през 1987 г. от златни водорасли. Той обединява монадични единични и колониални организми, понякога с редуване в жизнения цикъл на монадични и палмелоидни стадии. Повърхността на пектиновите черупки е покрита с обвивка от силициеви люспи. Митохондриите с тръбести кристи обикновено са разположени в цитоплазмата около хлоропласта. Ядрото е едно, хлоропластите са заобиколени от четири мембрани, обикновено две. Ламелите са тритилакоидни, има поясна ламела. Основните пигменти са хлорофилите АИ с, β -каротин и фукоксантин. Няма стигма. Клетките обикновено с 2 неравни флагела. Дългият перест камшик е насочен напред. Къс, гладък камшик, понякога силно намален, насочен назад.
Клетките се размножават главно чрез надлъжно делене. Колониите, разпадайки се, пораждат млади колонии. При някои видове половият процес е описан под формата на изогамия. В същото време, за разлика от златните водорасли, сливането на изогамети в синуричните водорасли се случва не в предните, а в техните задни краища. Силицираните цисти с пори се образуват ендогенно, подобно на тези на хризофитите.
Преобладаващо планктонни форми, палмелоидни стадии са част от бентоса. Повечето видове синурови водорасли са фототрофни сладководни организми.
Систематика.
Класът Sinura водорасли включва 7 разреда: Chloramoebales, Synurales, Rhizochloridales, Ochromonadales, Heterogloeales, Parmales и Thallochrysidales.
Разред Синура - Synurales. Монадичните форми, пектиновите клетъчни мембрани обикновено са покрити с черупка от силициеви люспи, циментирани с органична материя в един корпус. Хлоропласти един или два.
Род Синура(фиг. 38) - сладководни монадични форми с два неравни флагела и два париетални хлоропласта. Ядрото е с крушовидна форма, разположено в предната част на клетката. Зад ядрото има една голяма вакуола, а в задната част на клетката има няколко малки контрактилни вакуоли. Клетките са покрити с люспи, като плочки.Размножаването е най-често вегетативно, при някои е познат половият процес. Образува колонии (фиг. 39, А). Често кистите се появяват във всички клетки на колонията. С масово развитие Синураможе да придаде на водата неприятна миризма.

Ориз. 38. Външен вид на клетките на синура
Разред Ризохлоридни - Rhizochloridales. Предимно сладководни организми с ризоподиален тип диференциация на талуса, живеещи предимно в сладки водни тела.
Род Маломонас(фиг. 39, Б-Д) – едноклетъчна монада с един добре изразен флагел и един раздвоен хлоропласт.

Ориз. 39. Външен вид на синурични водорасли (по: G.A. Belyakova et al., 2006): А – деляща се колония Синура; Б, В – образуване на статоспори и Ж – външен вид Маломонас
Клетката е покрита с керемидени и спираловидни люспи, някои от тях носят игловидни шипове (фиг. 39, Ж). Характерно е наличието на статоспори (фиг. 39, Б, В). За редица представители е описан сексуалният процес, хологамия.
Род Миксохлорисживее в клетките на листата на сфагнови мъхове, има вид на голям многоядрен плазмодий. През есента образува цисти, които покълват през пролетта. От тях се появяват зооспори или амеба, които проникват в празни листни клетки и се сливат там в плазмодий.
Разред Chloramoebaceae - Chloramoebales.Включва представители на монадите. Среща се в солени и сладки води.
Род хетерохлорис – клетките му могат да променят формата си, образувайки псевдоподии. Това явление е присъщо на много представители на разреда, както и склонността към животинския начин на хранене. В клетката има няколко хлоропласта, маслени капки и хризоламинарин. Пред клетката има контрактилни вакуоли, в центъра - едно ядро. Размножава се вегетативно чрез клетъчно делене.
Разред Охромонадови - Ochromonadales. Съчетава голи форми с две неравни флагели. Сладководни и морски форми.
Род Охромонасвключва едноклетъчни монади с две неравни флагели. Клетките са облечени само от плазмалема. В предния край на клетката има контрактилна вакуола и око, в задния край има вакуола с хризоламинарин. Размножава се вегетативно, браздата на делене започва в предния край на клетката между две двойки флагеларни основи. Видовете от рода са често срещани в олиготрофните сладки води, но има и морски представители.
Разред Хетероглееви - Heterogloeales. Включва водорасли с палмелоиден тип диференциация на талуса. Представителите на разреда се срещат по-често в сладки, отколкото в солени води.
Род Хелминтоглеяживее в солени води. Тази колония, разположена върху разширена основа, се състои от разклонени лигави ленти. Протопластите са произволно подредени в тази слуз, всеки от които е заобиколен от собствена лигавична обвивка.
Клас Phaeotamnia водорасли - Phaeothamniophyceae
Името на класа произлиза от типа genus Феотамнион(от гръцки. phaeos- мургав храст). Представители на тази група организми бяха идентифицирани в отделен клас от жълто-зелени и златни водорасли през 1998 г. въз основа на анализа на последователността на рибозомните гени и особеностите на клетъчната ултраструктура. Водораслите Feotamnia се характеризират с уникална комбинация от пигменти: фукоксантин с хетероксантин, липса на виолаксантин.
Характеристики на клетъчната структура: липсата на вакуоли с хризоламинарин и липсата на ендогенни кисти със стени от силициев диоксид.
Класът включва едноклетъчни, колониални и многоклетъчни организми с кокоидна, палмелоидна и нишковидна структура на талуса. Камшичести стадии с две неравни камшичета. Камшичета странични или субапикални. Стигмата се среща в зооспорите. Пигменти - хлорофили АИ с, β -каротин, фукоксантин, диадиноксантин, диатоксантин и хетероксантин. Основният резервен продукт е парамилон ( β -глюкан). Клетъчната стена присъства, по време на клетъчното делене дъщерната стена се образува вътре в родителската стена.
При feotamniaceae от основните методи на размножаване се срещат само вегетативно и безполово. Безполовото размножаване се осъществява чрез автоспори или зооспори. Сексуалното размножаване не е известно.
Видовете водорасли Feotamnia живеят изключително в сладки води. Систематика.
Понастоящем е потвърден монофилетичният клас на водораслите феотамния, а сред охрофитите той е по-близо до кафявите и жълто-зелените водорасли, отколкото до златните водорасли. В момента са известни около 30 вида от тези водорасли, приписани на един разред Теотамния - Phaeothamniales.
Род Теотамнионпредставлява прикрепени разклонени нишки с височина до 1 cm (фиг. 40). Клетките са цилиндрични, неправилни, разширени нагоре, надлъжно

Ориз. 40. Външен вид Теотамнион.
два до три пъти по-дълъг от широк. По време на клетъчното делене материалът на родителската стена се изразходва за образуването на слоеста обвивка, обграждаща нишковидните клетки. По периферията на клетката под плазмената мембрана има везикули. Наподобяват физоди – образувания, намиращи се в клетките на кафявите водорасли. Клетките съдържат от един до няколко дисковидни маслиненокафяви пластиди. Зооспорите се образуват от 1–2, по-рядко от 4–8 в една клетка. При зооспорите флагелите са прикрепени странично. Положението на флагеларните корени Теотамнионнаподобява тази на жълто-зелените и кафявите водорасли. Липсва му и ризопласт. В жизнения цикъл могат да се образуват кисти, но в стените им няма силиций. Цистите покълват като зооспори. Заселва се като епифит върху нишковидни водорасли. Обитава стоящи и бавно течащи сладки водоеми.
Клас Rafid водорасли -Raphidophyceae
Името на класа идва от гръцки рафид - игла. Обединява едноклетъчни бифлагелатни организми, лишени от клетъчна стена. Предимно сладководни организми със сплескани клетки (фиг. 41).
Клетките имат флагели с различна дължина; камшикът, насочен напред, е перест и по-дълъг, камшикът, насочен назад, е гладък и по-къс. Преходната спирала липсва. Хлоропластите са малки, многобройни или по-рядко 1-2 на клетка, заобиколени от четири мембрани, от които две са CES мембрани. Тилакоидите са подредени в купчини по три. Ядрото е голямо, заобиколено от пръстен от диктиозоми. Митохондрии с тръбести кристи. Има контрактилни вакуоли. Цитоплазмата често е вакуолизирана. Клетките са голи, заобиколени само от плазмалемата. Стигма (око) обикновено отсъства.
Фотосинтетични пигменти – хлорофили АИ с, β -каротин, вошериаксантин, диноксантин, диадиноксантин, хетероксантин.
Резервни продукти за асимилация са мазнини и масла, по-рядко нишесте и хризоламинарин.
Те също живеят в сфагнови блата и други местообитания с кисела и неутрална водна реакция. Някои видове се срещат в солени и морски води.

Ориз. 41. Външен вид на рапидните водорасли.
Систематика.
Дълго време тази група водорасли, поради зеления си цвят, наличието на фаринкс и редица други структурни характеристики, се считаше за клас Chloromonas сред Euglenoids. Но данните от цитологията, биохимията, физиологията и молекулярната биология показаха безспорната им принадлежност към отдела на водораслите Heterocont (Ochrophyte) и монофилетичната природа на групата водорасли Rafid.
От самото са известни около 25 вида разред Huttonellaceae– Chattonellales.
Род хетеросигма(фиг. 42, А) включва морски флагелати. Клетки с овална форма с флагели, изместени настрани, пластидите са разположени по периферията на клетката. Няма трихоцисти. Те плуват, извършвайки въртеливи движения около надлъжната ос на тялото. Може да причини токсичен "цъфтеж на водата" в крайбрежните води.

Ориз. 42. Рафидни водорасли (от: R. E. Lee, 1999): А – хетеросигма; б – Гониостомум; 1 - хлоропласт; 2 - мукоцист; 3 - контрактилна вакуола; 4 - трихоцист; 5 - ядро
Род Гониостомум(фиг. 42, б) са подвижни дорзовентрално компресирани монади. Дорзалната страна е извита, коремната страна е сплескана. Една бразда в предния край води в триъгълен фаринкс, от който излизат две дълги камшичета; тяхната дължина е сравнима с дължината на клетката. Пластидите са разположени по периферията на клетката. Под плазмалемата има трихоцисти. Живее в сладки води с кисело pH.
Клас Eustigma водорасли -Eustigmatophyceae
Името на класа идва от гръцки ЕС- "добре и стигма- "белег", "петно". Класът обединява голи едноклетъчни, по-рядко колониални организми с предимно кокоидна структура.
Едно или повече ядра. Хлоропластът обикновено е един, заобиколен от 4 мембрани. Камшичето обикновено е едно предно пересто и второ базално тяло или по-рядко две неравни камшичета. Налице е стигма (око), разположена извън хлоропласта, нейните гранули са без мембрани.

Ориз. 43. Външен вид на водорасли Eustigma
Хлоропластът обикновено е единичен, голям, многолостен, чашовиден или париетален, жълто-зелен на цвят. Тилакоидите са събрани в ламели по три, обграждаща ламела липсва.
Основните пигменти са хлорофил а, β-каротин, виолаксантин и вошериаксантин. Пиреноидът обикновено присъства само във вегетативни форми.
Резервни вещества са масла и вещество с неизвестна природа, което се отлага като твърд материал извън пластидите. Нишестето не се образува.
Те живеят в сладки води, по-рядко се срещат в моретата и в почвата.
Систематика.
Преди това тази група организми е била класифицирана като жълто-зелени водорасли. В момента водораслите Eustigma се считат за ранг на клас в отдела на Heterocontophytes. Известни са около 35 вида, предимно сладководни, приписвани на единствения Разред Еустигма - Eustigmatales.
Род Евстигматос(фиг. 43) - едноклетъчни малки сферични водорасли. Килийната стена е гладка, цяла, без орнаменти. Жълто-зелен хлоропласт, единичен, дялан, теменен, с дръжкови полиедрични пиреноиди. Голяма централна вакуола с червено съдържание. Размножава се от 2 или 4 автоспори или зооспори във формата на бутилка с един входящ камшик, голяма стигма, разположена извън хлоропласта в предния край, и единичен заден хлоропласт без пиреноид. Често срещан компонент на почвената флора. Известно от Нова Зеландия, Австрия, Исландия и от басейните на Аризона като "синапено водорасло".
Клас Жълто-зелени водорасли -Xanthophyceae
Жълтозелените водорасли са тези, чиито хлоропласти са оцветени в светло или тъмно жълто, много рядко в зелено и само понякога в синьо. Цветът на талиите се определя от наличието на следните пигменти в клетките на хлоропластите - хлорофили АИ с, β -каротин и ксантофили. Преобладаването на последното определя оригиналността на цвета на жълто-зелените водорасли. В допълнение, парамилон, маслени капки и само при някои видове, в допълнение, бучки от левкозин и волутин се натрупват в клетките като основен продукт на асимилацията. Жълто-зелените водорасли не образуват нишесте. Отличителна черта на жълто-зеленото е наличието на монадична структура във вегетативните клетки и две неравни флагели в зооспорите. Клетъчната стена съдържа целулоза, глюкоза и уронова киселина. Клетъчната стена често се състои от две части.
Размножаването е вегетативно, безполово и полово.
Широко разпространен в сладки води. Рядко се среща в морски, солени води и в почвата.
Преди това класът жълто-зелени водорасли се е наричал водорасли Tribophytium, след вида на рода Tribonema (от гръцки. трибон – умел, хитър и няма – нишка). Известни са около 450 вида.
Жълтозелените се характеризират със значително морфологично разнообразие. Сред многобройните представители на този отдел се срещат почти всички основни типове структура на тялото: амебоидна, монадична, палмелоидна, кокоидна, нишковидна, многонишкова, ламеларна и сифонална (фиг. 44). – 46). Талус едноклетъчен,

Ориз. 44. Външен вид на жълто-зелени водорасли: 1, 2 - Харатиопсис, 3 – Centritractus, 4 – офиоцитиум
колониални, многоклетъчни и неклетъчни. Клетъчната мембрана е плътна, пектинова и целулозна, състояща се от плътно припокриващи се части или от две клапи. В черупката се отлага силициев диоксид или вар. Предимно фиксирани форми. Сред едноклетъчните видове има подвижни форми, лишени от плътна обвивка и оборудвани с флагели, лобоподии и кореноподии.

Ориз. 45. Външен вид на ксантококови жълто-зелени водорасли: 1–3 - Ботридиопсис, 4 – тетраедриела, 5 – Pseudostaurastrum, 6 – Гониохлорис, 7, 8 – Бумилериопсис
Повечето жълто-зелени – неподвижни организми. При подвижните индивиди движението може да се извършва с помощта на жгутици или ризоподии. Клетки с различни форми: сферични, вретеновидни, елипсовидни, цилиндрични, тетраедрични, сърповидни, крушовидни, яйцевидни. Размери на талуса от 0,5 – 1,5 µm ( Хлоридела) до няколко милиметра в диаметър ( Ботридиопсис) (фиг. 45, 1 – 3) и до десетки сантиметри дължина ( Вошерия) (фиг. 46, 3).

Ориз. 46. Външен вид на жълто-зелени водорасли: 1 - Трибонема, 2 – хетеропедия, 3 – Вошерия, част от нишка с оогоний и антеридий
Повечето жълто-зелени видове са фототрофи, но се открива и холозойно хранене чрез поглъщане на бактерии и малки водорасли. Жълто-зелените водорасли са широко разпространени в сладките води. Те също са често срещани в почвата, по-рядко в морските и солените води. Класът включва аеробионтни, планктонни, бентосни и перифитонни форми. Епифити, епизоити, както и вътреклетъчни симбионти в протозойни клетки.
Независимо от външната структура, вътрешната структура на клетката на жълто-зелените водорасли е една и съща. В протопласта обикновено има няколко жълто-зелени хлоропласта, които имат дисковидна, коритовидна, ламеларна, по-рядко лентовидна, звездовидна или чашковидна форма с цели или лобовидни ръбове. Оцветяването се дължи на липсата на фукоксантин, който е отговорен за златния и кафяв цвят при други охрофити. От другите пигменти, които имат β -каротин, вошериаксантин, диатоксантин, диадиноксантин, хетероксантин. При подвижните форми червено око или стигма обикновено се намира в предния край на хлоропласта. Малко видове имат пиреноиди от полупотопен тип. Ядрото в клетката обикновено е едно, малко по размер, но има видове с многоядрени клетки. Някои видове имат една или две контрактилни (пулсиращи) вакуоли в предната част на клетката.
Представителите на монадите и подвижните стадии (зооспори и гамети) имат два неравни флагела. Изключение правят синзооспорите. Вошери, в който по повърхността са разположени многобройни двойки гладки камшичета, леко различни по дължина. Къс флагелум завършва с акронем. Флагелите са прикрепени към клетката субапикално. При сперматозоида Вошеристранично закрепване.
При видовете с амебоидна, монадална и палмелоидна организация клетъчната стена отсъства, те са покрити само от цитоплазмена мембрана и могат лесно да променят формата си. Понякога "голи" клетки се намират вътре в къщи, чиито стени могат да бъдат боядисани в кафяво с манганови и железни соли. По-голямата част от формите имат клетъчна стена, състояща се от две части. Целулозата преобладава в състава на клетъчната стена, а също така съдържа полизахариди, състоящи се главно от глюкоза и уронова киселина. В младите клетки мембраната е тънка, с възрастта се удебелява. В него могат да се отлагат железни соли, чиито съединения го оцветяват в различни нюанси на кафяво и червено. По-често силициевият диоксид присъства в клетъчната стена, което й придава твърдост и блясък. Може също така да бъде инкрустиран с вар и да бъде изваян по различни начини (бодли, клетки, брадавици, черупки, зъбци и др.) Израстък на черупката може да се образува в прикрепени форми – крачол с подметка за прикрепване.
При нишковидните форми на жълто-зелени водорасли с двучерупчести черупки, когато нишките се разпадат, клетъчните мембрани се разпадат на Н-образни фрагменти. Тези фрагменти са плътно свързани половини на черупките на две съседни клетки (фиг. 47). По време на растежа на нишките, Н-образен фрагмент от клетъчната стена на две съседни дъщерни клетки се вгражда между двете половини на мембраната на майчината клетка. В резултат на това всяка от дъщерните клетки е наполовина покрита със старата мембрана на майчината клетка и наполовина – новообразувана черупка.

Ориз. 47. Схема на образуване на напречна преграда между две дъщерни клетки в нишковидни жълто-зелени водорасли (според: A.A. Masyuk, 1993): А- фрагмент от нишка; б- полагане на поясния пръстен и образуване на напречна преграда между две клетки; IN– наслояване на двучерупчести клетъчни мембрани; Ж– разпадане на черупката на Н-образни участъци
Съкратителните вакуоли присъстват в подвижните представители. Обикновено има 1-2 на клетка. Апарат на Голджи със специфична структура. Диктиозомите са малки, съдържат 3-7 цистерни. Ядрото е едно, по-рядко са много; при ценотичните видове клетките винаги са многоядрени.
размножаване. Повечето видове жълто-зелени водорасли се характеризират с вегетативно и безполово размножаване.
Вегетативно размножаванеизвършва се по различни начини: клетъчно делене наполовина, разпадане на колонии и многоклетъчни тали на части. При Вошериобразуват се специални плодови пъпки.
При безполово размножаванеМогат да се образуват различни спори: амебоиди, зооспори, синзооспори, автоспори, хемизооспори, полуавтоспори, апланоспори. Зооспорите са "голи" и обикновено имат крушовидна форма.
полов процес- изогамия, хетерогамия и оогамия - описани в няколко представителя. При Трибонемигаметите са сходни по размер, но се различават по поведение – това е изогамия. При Вошеринаблюдава се оогамия: върху нишките се образуват вместилища на женски гамети – оогония и мъжки – антеридии.
При неблагоприятни условия се наблюдава образуване на кисти. Цистите (статоспорите) са ендогенни, едноядрени, рядко многоядрени. Стената им често съдържа силициев диоксид и се състои от две неравни или по-рядко равни части.
Систематика.
В края на XIX – началото на 20 век различни родове жълто-зелени водорасли бяха причислени към отдела на зелените водорасли, което беше свързано предимно с цвета и морфологичното сходство на талиите. Жълтозелените понастоящем се считат за клас в рамките на охрофитния отдел.
Известни са около 450 съвременни вида от клас жълто-зелени водорасли, които са групирани в четири разреда: Botridia, Mishococcus, Tribonem и Vosheria. Класификацията на разредите се основава на вида на диференциацията на талуса и характеристиките на жизнения цикъл.
Разред Ботридиеви - Botrydiales. Разредът включва видове със сифонален тип диференциация на талуса, при които липсва оогамен полов процес.
Род Ботридиумживее върху почвата и има вид на зелени мехурчета с размери няколко милиметра, прикрепени с безцветни ризоиди. Талусът е сифонален, съдържа множество ядра и пластиди. Черупката е многопластова, върху нея може да се отлага вар. Размножаването е безполово с помощта на бифлагелатни зооспори, докато цялото съдържание на пикочния мехур се разпада на едноядрени фрагменти. При липса на влага се възпроизвежда с помощта на апланоспори или образува дебелостенни кисти. В някои случаи цялото съдържание на пикочния мехур отива до образуването на една голяма киста. В други случаи кистите се образуват в ризоидите, където първо преминава съдържанието на пикочния мехур. Цистите покълват или директно в новия талус, или образуват зооспори. Половият процес е изо- и хетерогамия. Зиготата покълва веднага, без период на покой. Често срещан и широко разпространен вид в сухоземните местообитания, намиращ се по бреговете на потоци, езера или върху почви без растителност.
Поръчайте Michococcal–Mishococcales. Едноклетъчни или колониални представители с кокоиден тип диференциация на талуса.
Род Харатиопсисвключва едноклетъчни прикрепени форми. При размножаване образува зооспори, апланоспори и дебелостенни цисти (фиг. 44, 1-2).
Род офиоцитиум(фиг. 44, 4) има продълговати цилиндрични килийки, които могат да бъдат прави, огънати или спирално усукани, като на края могат да носят шип. Клетъчната стена се състои от две неравни части, от които по-голямата част участва в растежа на клетките, по-малката част е постоянна и има формата на шапка. Едноклетъчни и колониални видове, свободно живеещи или прикрепени към субстрата с малка дръжка. Размножават се чрез зооспори и апланоспори, срещат се цисти. Те живеят в сладки води.
Род Mischococcusобразува дървовидно прикрепени колонии. Разклоняването е дихотомно и тетрахотомно. Клетките са разположени по 2 или 4 по върховете на лигавичните разклонения на колонията. Клетките са сферични до овални, с тънка или дебела клетъчна стена. Понякога клетъчната стена е лъскава и кафява поради импрегнирането й с железни соли. Млади едноклетъчни организми със слузеста дисковидна основа, която служи като опорна точка за прикрепване. След освобождаването на спорите протопластът на майчината клетка се превръща в желе и се разтяга, дължината става 6 пъти по-голяма от ширината и така се появява цилиндричен крак. Празната клетъчна стена на майчината клетка винаги става основата на стъблото. Безполово размножаване чрез зооспори и автоспори. Автоспорите са прикрепени към горния ръб на слизестата дръжка. Последващите клетъчни деления повтарят процеса и произвеждат дървоподобна колония. полов процес – изогамия. Те живеят в малки сладки водоеми като епифити на нишковидни водорасли. Известен в Централна Европа и Азия.
Разред Tribonemaceae - Tribonematales. Представителите имат нишковиден, мултифиламентозен, фалшивотъканен и тъканен тип диференциация на талуса. Клетъчните стени са или с H-образни припокриващи се части, или са твърди.
Род Трибонема- неразклонени нишки (фиг. 46, 1). Клетките са цилиндрични или бъчвовидни. Клетъчната стена се състои от две половини, които се намират с ръбове един върху друг в средата на клетката. Черупките често са наслоени. Фрагменти от нишки винаги завършват с празни половини на H-образни фрагменти от черупки, които приличат на вилица. В клетките има няколко жълтеникаво-зелени пластиди, няма пиреноиди. Размножаването е вегетативно (фрагментиране на нишките), безполово (чрез зооспори и апланоспори) и полов път (изогамия), като апланоспорите се произвеждат по-често от зооспорите. Може да образува акинет. Те живеят в сладки води, където се развиват особено изобилно през студения сезон.
Разред Vosherievye - Vaucheriales. Всички представители имат сифонален талус, оогамен полов процес и синзооспори.
Род Вошерия(фиг. 46, 3) има талус с неклетъчна структура; неговият талус достига дължина от няколко сантиметра, прикрепен към субстрата с помощта на безцветен ризоид. В нишките няма прегради, по-голямата част от талуса е заета от вакуола, а множество ядра и пластиди са разположени по периферията в цитоплазмата. Нишки с апикален растеж и рядко странично разклонение. Преградите се образуват, когато талусът е повреден и за отделяне на репродуктивните органи. Безполовото размножаване се осъществява от апланоспори, синзооспори, акинети. Синзооспорите се образуват една по една в зооспорангиума, който е отделен от вегетативните клетки чрез преграда в края на нишката. Зооспорите са многоядрени и многофлагелирани. Половият процес е оогамия. Зиготата е покрита с дебела обвивка и след период на покой прераства в нов талус.
Видове Вошеришироко разпространен в сладки, солени и морски води, както и в сухоземни местообитания. Срещат се на всички континенти, включително Антарктида. Те образуват тревисто-зелени или тъмнозелени преплетени маси - така наречените рогозки, гладки, пълзящи или възглавничести. Водни, полуводни, сухоземни форми. Те живеят в различни местообитания: морета, естуари, устия, солени блата, мангрови гори, потоци, канали, езера, езера, обработваема земя и блата.
Значениехетероконтови водорасли
От отдела за водорасли Heterocont, кафявите водорасли са от най-голямо значение за природните екосистеми и за хората.
Кафяви водорасли - осн източник на органична материяв крайбрежната зона на моретата. Тяхната биомаса в моретата на умерения и субполярния пояс може да достигне няколко десетки килограма на квадратен метър. Гъсталаците от кафяви водорасли осигуряват условия за хранене и размножаване на много крайбрежни животни и други водорасли. Чарлз Дарвин наблюдава кафяви водорасли край бреговете на Южна Америка Макроцистис, написа: „Мога само да сравня тези обширни подводни гори на южното полукълбо със сухоземните гори на тропическите региони. И все пак, ако една гора бъде унищожена в някоя страна, не мисля, че дори приблизително същият брой животински видове ще умрат, както при унищожаването на тези водорасли.
Служат гъсталаци от кафяви водорасли място за храна,подслон и отглежданемного животни. Образно казано, кафявите водорасли осигуряват на другите водни организми „маса, подслон и детска стая“.
Кафявите водорасли се използват широко от хората. Те са богати йоди други микроелементи. Народите от Югоизточна Азия традиционно ги използват за храна, особено представители на разред водорасли, от които се приготвят голямо разнообразие от ястия. Фуражно брашно, приготвен от кафяви водорасли, повишава продуктивността на добитъка; в същото време се увеличава съдържанието на йод в яйцата и млякото.
От кафяви водорасли получавамалгинати- соли на алгиновата киселина. Алгинатите се използват широко в различни индустрии. Това са нетоксични съединения с колоидни свойства, така че се използват широко в хранително-вкусовата и фармацевтичната промишленост. Алгиновата киселина и нейните соли са способни на 200-300-кратна абсорбция на вода, образувайки гелове, които се характеризират с висока киселинна устойчивост. В хранително-вкусовата промишленост те се използват главно като емулгатори, стабилизатори, желиращи и водозадържащи компоненти. Например, сух прах натриев алгинат се използва в производството на прахообразни и брикетирани разтворими продукти (кафе, чай, мляко на прах, желе и др.) За бързото им разтваряне. Водните разтвори на алгинати се използват за замразяване на месни и рибни продукти. До 30% от общия обем алгинати, произведени в света, отиват в хранително-вкусовата промишленост.
В текстилната и целулозно-хартиената промишленост алгинатите се използват за сгъстяване на бои и увеличаване на здравината на връзката им с основата. Импрегнирането на тъканите с някои алгинати им придава защитни свойства: водоустойчивост, киселинна устойчивост и повишава механичната якост. Редица соли на алгиновите киселини се използват за получаване на изкуствена коприна. По време на Втората световна война в САЩ и Англия от алгинова киселина и нейните соли се произвежда голямо количество камуфлажни тъкани и мрежи за жилищни и промишлени сгради.
Алгинатите се използват в металургията: в леярната те подобряват качеството на формовъчната пръст. Солите на алгиновата киселина се използват при производството на електроди за електрозаваряване, което позволява получаването на по-високо качество на заварките. Алгинатите се използват и в производството на пластмаси, синтетични влакна, покрития и устойчиви на атмосферни влияния строителни материали. Използват се при производството на висококачествени смазочни материали за машини. В радиоелектрониката алгинатите действат като свързващ агент при производството на висококачествени ферити.
Най-голямо приложение намира водоразтворимият натриев алгинат, способен да образува вискозни разтвори. Използва се широко за стабилизиране на различни разтвори и суспензии. Добавянето на малко количество натриев алгинат към хранителни продукти - консерви, сладолед - подобрява тяхното качество. Използва се и за направата на декоративна козметика, кремове и маски в парфюмерийната индустрия.
Във фармацевтичната индустрия алгинатите се използват за покриване на таблетки, хапчета, като компонентни основи за различни мехлеми и пасти, като гелове за носители на лекарства и при производството на разтворими хирургически конци. В медицината калциевият алгинат се използва като хемостатично средство и като сорбент, който премахва радионуклидите (например стронций). Годишното производство на алгинати в света надхвърля 20 хиляди тона.
Друго важно вещество, получено от кафяви водорасли, е шестводният алкохол манитол. Манитолът се използва като заместител на захарта при диабетици. Освен това може да се използва като плазмозаместител за консервиране на кръвта. Използва се за производство на таблетки във фармацевтичната индустрия. Манитолът се използва и при производство на синтетични смоли, бои, хартия, експлозиви, обработка на кожи.
фукоидани,получени от кафяви водорасли са ефективни антикоагуланти, дори по-активни от хепарина. Използването им за производството на противотуморни лекарства и антивирусни съединения се счита за обещаващо. В края на краищата, дори при най-ниски концентрации, фукоиданите могат да инхибират прикрепването на вирусите към повърхността на клетките. Фукоиданите също имат способността да образуват изключително силна и вискозна слуз, която се използва при приготвянето на стабилни емулсии и суспензии.
Енергийната криза, която обхвана много страни по света през последните години, доведе до необходимостта от търсене на нови нетрадиционни източници на енергия. И така, в САЩ за тази цел се проучва възможността за отглеждане на гигантски водорасли водорасли. Макроцистиспоследвано от преработката му в метан. Изчислено е, че от площ от 400 km 2, заета от това водорасло, могат да се получат 620 милиона m 3 метан.
Хетероконтните водорасли от класовете златни, жълто-зелени, синура, рафидофитни и евстигма водорасли, представени главно от микроскопични организми, са широко разпространени в сладките водоеми на всички климатични зони на земното кълбо, но са по-често срещани в умерените ширини. Сред златните водорасли има видове, които живеят в моретата и солените езера и много малко в замърсените води. Златните водорасли достигат максимално развитие през студения сезон: те доминират в планктона през ранна пролет, късна есен и зима. По това време те играят значителна роля като производители на първична продукция и служат като храна за зоопланктонните организми.
Някои златни водорасли, например УрогленаИ Динобрион,развивайки се в масови количества, те са в състояние да предизвикат цъфтеж на водата. Те отделят алдехиди и кетони, които могат да придадат на водата неприятна миризма и вкус, а Уроглена– мастни киселини, токсични за рибите.
Рафидните водорасли са широко представени в планктона на сладководни тела с преобладаващо киселинно рН, особено в сфагнови блата, по-рядко в големи езера. В сладки водни тела могат да се образуват местни "цъфтежи". Гониостомум.Рафидните водорасли се срещат и в обезсолени морски заливи и локви на морския бряг, както и в открито море. С масово развитие в крайбрежните морски води те предизвикват токсично „цъфтене” на водата. И така, край бреговете на Канада по време на "цъфтежа" концентрацията на клетките на рафидното водорасло Heterosigma може да достигне 30 милиона на 1 литър. Избухването на рафидни водорасли често води до развитие на "червени приливи", които са свързани със смъртта на рибата. Причината за такива "червени приливи" може да бъде вид раждане Hattonella, Olistodiscus, Heterosigma и Fibrocapsa.
Водораслите Sinura с масово развитие в сладки водоеми могат да придадат на водата неприятна миризма ( Синура). Водораслите Feotamnia се срещат в застояли и бавно течащи сладки водоеми, където се установяват епифитно върху нишковидни водорасли.
Водораслите Eustigma се срещат само в прясна вода или в почвата.
Жълто-зелените водорасли са разпространени на всички континенти, живеят предимно в сладки води и почви, както и в сухоземни, соленоводни и морски местообитания. Жълто-зелените водорасли обитават чисти и замърсени води с различни стойности на pH: те могат да живеят както в кисели, така и в алкални води. Те се срещат главно в чисти сладководни резервоари, по-рядко в морета и солени води, предпочитайки умерени температури, по-често се развиват през пролетта и есента, въпреки че има видове, които се срещат през всички периоди на годината, включително зимата. Най-често те могат да бъдат намерени в групи от нишки и сред гъсталаци на висши водни растения в крайбрежната зона на реки, езера, езера и резервоари.
По-голямата част от жълто-зелените са свободно живеещи форми, но има и вътреклетъчни симбионти - зооксантели в протозойни клетки. Интересна вътреклетъчна симбиоза се образува от хлоропластите на морските видове Вошерис мида Елизион. В продължение на девет месеца това мекотело е способно на фотоавтотрофно фиксиране на въглероден диоксид в културата. Това е най-дългата симбиоза от този тип, когато симбиотичният пластид е в пряк контакт с цитоплазмата на животното. В природата ларвите на мекотелите се хранят с нишки. Вошери. В резултат на фагоцитозата хлоропластите на водораслите навлизат в цитоплазмата на епителните клетки на мекотелите. По време на този процес хлоропластната мембрана става трислойна, една външна мембрана на CES (хлоропластен ендоплазмен ретикулум) се губи. Това явление е добро доказателство, че в хода на еволюцията хлоропластите с три мембрани могат да възникнат в резултат на вторична симбиогенеза поради загуба на мембрани.
Жълтозелените, златните и други хетероконтни водорасли са производители на кислород и органични вещества; те са част от хранителната верига. Хетероконтофитите участват в самопречистването на замърсени води и почви, образуването на сапропел, в процеса на натрупване на органични вещества в почвата, оказвайки влияние върху нейното плодородие. Те се използват като представителни организми при определяне на състоянието на замърсяване на водата; жълто-зелените водорасли са част от комплекс от микроорганизми, използвани за пречистване на отпадъчни води.
Контролни въпроси
Назовете характерните черти на структурата на кафявите водорасли.
Структурни особености на талиите на кафявите водорасли.
Как се размножават кафявите водорасли? Какво представляват моноспорите, тетраспорите и зооспорите, изогамията, хетерогамията и оогамията?
Какви са жизнените цикли на кафявите водорасли? Размножаване на водорасли фукус и келп.
Назовете характерните особености и типичните представители на разредите кафяви водорасли.
В какви местообитания се срещат кафявите водорасли? Какво е значението им в природата.
Стопанско значение на кафявите водорасли.
Назовете характерните черти на структурата и типичните представители на златните водорасли.
Какви пигменти и видове хранене са известни в златните водорасли?
Размножаване и екология на златните водорасли.
Назовете характеристиките и типичните представители на жълто-зелените водорасли.
Какви пигменти и видове хранене са известни в жълто-зелените водорасли?
Как се размножават жълто-зелените? Видове сексуално размножаване: изогамия, хетерогамия и оогамия?
Назовете характерните особености на структурата и типичните представители на синуричните водорасли.
Размножаване и екология на синуричните водорасли.
Назовете характерните особености на структурата и типичните представители на водораслите феотамния.
Размножаване и екология на феотамниевите водорасли.
Назовете общите характеристики на структурата и типичните представители на рафидофитните водорасли.
Размножаване и екология на рафидофитни водорасли.
Кои са общите черти в структурата и типичните представители на водораслите Eustigma.
Размножаване и екология на водораслите Eustigma.
Стойността на хетероконтните водорасли в естествените екосистеми.
Flagella - най-старата група протозои, лежаща в основата на генеалогичното дърво на животинския свят и свързваща животните с растителния свят. Още в средата на предишния век известният руски биолог Л. С. Ценковски (1822-1887) посочи липсата на рязка граница между флагелираните протозои и едноклетъчните водорасли. В действителност, някои от камшичестите (еугленоиди и фитомонади) са основателно класифицирани както като тип протозои, т.е. животни, така и като тип зелени водорасли, т.е. растения.
Характеристика на класа
Една от най-важните характеристики, използвани при класификацията на протозоите, са органелите на движението. При флагелатите това са снопчета или бичове - тънки цитоплазмени израстъци. Те обикновено са разположени в предния край на тялото и възникват от специални базални зърна (базално тяло или кинетозома). Броят на камшичетата при различните видове варира от 1 до 8 или повече. В някои флагели (лейшмания, трипанозоми), в основата на флагела, освен това се поставя специален органоид, кинетопласт. По своята ултраструктура съответства на митохондриите, но се отличава с високо съдържание на ДНК. Смята се, че в кинетопласта се генерира енергия за движението на флагела, който извършва въртеливо движение и сякаш се завинтва във водата. При някои представители на класа флагелът минава по тялото, свързвайки се с него с тънък израстък на цитоплазмата. Споменатият израстък, или вълнообразна мембрана, извършва вълнообразни движения и служи като допълнителен органел на движение.
Според начина на хранене (усвояване) се делят на:
Размножаването обикновено е безполово, чрез надлъжно разделяне на две части. Много видове имат и полов процес, при който половите форми се сливат (копулация).
В застояли сладководни резервоари живеят многобройни видове зелени биченоси (euglena и phytomonads). Понякога се появяват в толкова големи количества, че водата става зелена (цъфтеж на водата).
Самотни свободно живеещи форми
Типичен представител на единичните форми е зелената еуглена (Euglena viridis). Формата на тялото му е вретеновидна, постоянна поради уплътняването на външния слой на протоплазмата. В цитоплазмата, в задната част на тялото, има голямо сферично ядро.
В предния край на тялото на зелената еуглена има тънък флагел, поради въртенето на който се движи. Има и пулсираща вакуола, нейният резервоар и няколко съдържащи хлорофил хроматофори, в които въглехидратите се синтезират от въглероден диоксид и вода (фотосинтеза) на светлина, т.е. наблюдава се автотрофно хранене. Продуктите от фотосинтезата се отлагат под формата на нишестеподобно вещество - парамил. Еуглената търси осветени места с помощта на светлочувствително око - фоторецептор с червен цвят, също разположен в предния край на тялото.
Euglena може да се храни и по холозойски начин, поглъщайки оформени хранителни частици. Ако околната среда е богата на разтворени органична материя, след което чрез осмоза навлизат в тялото на еуглената. По този начин в структурата и начина на хранене на еуглена се отбелязва комбинация от свойствата на животински и растителен организъм.
Поддържането на определена концентрация на соли в цитоплазмата, т.е. осмотичното налягане, се осъществява от специален органоид - пулсираща или контрактилна вакуола, разположена в предния край на тялото. Пулсиращата вакуола периодично увеличава обема си. Достигнал определен размер, той излива натрупаната течност през резервоара и след известно време започва да расте отново. Течността навлиза във вакуолата през аддукторните канали.
Размножаването на еуглената е безполово, чрез надлъжно делене на тялото. Първо ядрото се дели, базалното тяло, хроматофорите се удвояват и след това цитоплазмата се дели. Флагелумът се изхвърля или преминава към един индивид, а при друг се формира наново. Разделянето в надлъжна посока е едно от характерни особеностиклас флагели. При неблагоприятни условия настъпва енцистация: камшичето се прибира, еуглената се закръгля и образува плътна черупка. Инцистираната еуглена може да се дели един или повече пъти, докато остава вътре в черупките.
Различните видове еуглена са характерни за водни обекти с различна степен на органично замърсяване, следователно, заедно с водораслите, те могат да служат като критерий за санитарна оценка на източниците на водоснабдяване.
Колониални форми

Колониалните флагелати (волвокс, пандорина, еудорина и др.) се считат за преходни форми от едноклетъчни към многоклетъчни организми. Най-просто подредените колонии се състоят от 4-16 абсолютно идентични едноклетъчни индивида, свързани заедно - зооиди. Всеки зооид има флагелум, оцелус, хроматофори и контрактилна вакуола.
Представителят на колониалния вид флагелати - волвоксът (Volvox globator) образува големи сферични колонии, състоящи се от много хиляди вегетативни зооиди - малки крушовидни клетки, всяка от които има две камшичета. Диаметърът на топката е 1-2 мм. Кухината му е изпълнена с желатинообразно вещество. Всички клетки на Volvox (зооиди) са свързани помежду си с тънки протоплазмени мостове, което прави възможно координирането на движението на флагелата. Колонията се движи във водата поради координираното движение на камшичетата на отделните индивиди.
При Volvox вече се наблюдава разделение на функцията на клетките на колонията. И така, на единия полюс на колонията, по който тя се движи напред, има клетки с по-развити светлочувствителни очи, а в долната част на колонията (където очите са слабо развити) има клетки, способни да се делят (репродуктивни клетки, генеративни зооиди), т.е. отбелязва се диференциация на соматични и сексуални индивиди.
Възпроизвеждането на Volvox се извършва за сметка на специални - генеративни - зооиди. Те напускат повърхността вътре в колониите и тук, размножавайки се чрез делене, образуват дъщерни колонии. След смъртта на майчината колония, дъщерните започват самостоятелен живот. През есента поради генеративни индивиди се образуват и сексуални форми: големи неподвижни макрогамети (женски репродуктивни зооиди) и малки микрогамети, снабдени с две въжета (мъжки репродуктивни зооиди). В процеса на гаметогенеза индивидите, които се превръщат в макрогамети, не се делят и се увеличават по размер. Индивидите, които дават микрогамети, се делят многократно и се образуват голямо числодребни бифлагелати индивиди. Микрогаметите активно търсят неподвижни макрогамети и се сливат с тях, за да образуват зиготи. Зиготите пораждат нови колонии. Първите две деления на зиготата са мейотични. Следователно при колониалните флагелати само зиготата има диплоиден набор от хромозоми, всички останали етапи от жизнения цикъл са хаплоидни.
Подреждане на първични монади (Protomonadina) Семейство Трипанозоми (Trypanosomatidae) Род Leishmania (Leishmania)
Лайшмания - причинители на лайшманиоза - трансмисивни болести с естествена огнище. Лейшманиите са открити от руския лекар П. Ф. Боровски през 1898 г. Най-голямо значение имат представителите на рода Leishmania, принадлежащи към семейство Trypanosome, чиято отличителна черта е способността да образуват няколко морфологично различни формив зависимост от условията на съществуване. Промяната на формата се случва както при безгръбначни, така и при гръбначни гостоприемници.


Има следните морфологични форми:
- трипанозомна форма - характеризира се със сплескано лентовидно тяло, в центъра на което има овално ядро. Камшичето започва зад ядрото. Аксиалната нишка на флагела отива към предния край на тялото, образувайки добре развита вълнообразна мембрана. В предния край на тялото той завършва и флагелът стърчи напред, образувайки дълъг свободен край.
- Критидиална форма - камшикът започва малко по-напред от ядрото, придвижвайки се напред, образувайки къса вълнообразна мембрана и свободен край.
- leptomonas форма - флагелът започва от самия ръб на предния край на тялото, няма вълнообразна мембрана, свободният край на флагела е със значителна дължина.
- лейшманиална форма - има заоблена форма и голямо кръгло ядро. Пръчковидният кинетопласт е разположен в предния край на тялото. Флагелумът или отсъства, или има само вътреклетъчната му част, не излиза извън тялото.
- метациклична форма - подобна на критидиалната, но лишена от свободен флагел. (Фиг. 1).
Камшичестите от рода Leishmania имат две морфологични форми - лептомонас и лейшманиал (фиг. 2) или вътреклетъчни.
Лайшманията се разделя на дерматотропни (разпространени в кожата) и висцеротропни видове (разпространени във вътрешните органи).
Причинителят на висцералната лайшманиоза (Leischmania donovani, L. infantum)
Локализация. Клетки на черния дроб, далака, костния мозък, лимфните възли, ретикулоендотелните клетки на подкожната тъкан.
. Висцералната лайшманиоза е най-често срещана в Индия (патоген L. donovani), среща се в средиземноморските страни, в Закавказието и Централна Азия (патоген L. infantum), където се нарича кала-азар. В няколко страни в Азия, Африка и Южна АмерикаБолестите от лайшманиоза се причиняват и от други видове лайшмания.
. Лептомонадни и лайшманиални форми.

IN напоследъкУстановено е също, че Leishmania се намира в ретикулоендотелните клетки на кожата, което обяснява начина на заразяване на комарите. Засегнатите клетки понякога образуват непрекъснат слой или се концентрират в близост до потните жлези и съдове.
Патогенно действие. Има нередовна, постоянна треска. Далакът и черният дроб постепенно се увеличават и могат да достигнат огромни размери (фиг. 3). Развива се изтощение, съдържанието на еритроцити в кръвта намалява, възниква анемия. Заболяването може да бъде остро или да има хроничен курс (1-3 години). Смъртността е много висока. Боледуват предимно деца.
Предотвратяване: лични - индивидуална защита срещу ухапване от комари; обществен - набор от мерки за борба с комари и гризачи, унищожаване на бездомни и лайшманиозни кучета, чакали. В същото време е необходимо да се извършва санитарно-просветна работа и лечение на пациенти.
Кожна лайшманиоза (Leishmania tropica)
Известни са три подвида на дерматотропния вид Leishmania: L. tropica minor и L. tropica major (в източното полукълбо) и L. tropica mexicana (в западното полукълбо).
Географско разпространение. Разпространен в редица страни в Европа, Азия, Америка, Африка. В Русия кожната лейшманиоза, причинена от L. tropica (болест на Боровски), е унищожена, докато L. major се среща в Закавказието и Централна Азия, особено в редица региони на Туркменистан.
Морфофизиологични характеристики. Лептомонасната и лайшманиалната форма са неразличими от формите на висцеротропната лайшмания.
Жизнен цикъл. Почти не се различава от причинителя на висцералната лейшманиоза. Източник на инфекцията са хора и диви животни (малки гризачи, живеещи в зоните на пясъчните полупустини и пустини - песчанки, земни катерици, хамстери, някои видове плъхове и мишки). Заразеността на резервоарните животни в естествени условия понякога достига 70%. Заболяването при животните също се проявява под формата на кожни язви. Комарите служат като преносители. Съществува близък контакт между резервоарните гризачи и комарите-преносители. Дупката на гризачите е постоянно местообитание и място за размножаване на комари, чиято заразеност може да достигне 35%. Ролята на човека в разпространението на кожната лайшманиоза е малка, с изключение на някои райони на земното кълбо (Индия).
Патогенно действие. При кожна лайшманиоза се образуват закръглени, дълги (около година) незаздравяващи язви по отворени частитялото, особено по лицето. След зарастване остава обезобразяващ белег (фиг. 4). След заболяване имунитетът остава за цял живот.
Лабораторна диагностика. Микроскопско изследване на язви от отделяне.
Предотвратяване: лични - индивидуална защита срещу ухапване от комари; препоръчително е да се извършват профилактични ваксинации на щамове на кожна лейшманиоза от животно в затворени участъци от кожата. Такива ваксинации допълнително предпазват от появата на обезобразяващи язви и белези по лицето и други открити части на тялото. Като обществена профилактика се води борба с комари и гризачи. По-специално, те въвеждат хлорпикрин в дупките, разорават парцели земя, обитавани от гербили, в териториите, съседни на селата.

Род Trypanosoma
Trypanosoma gambienseе причинителят на трипанозомозата, африканската сънна болест при хората.
Локализация. В тялото на човека и други гръбначни животни той живее в кръвната плазма, лимфата, лимфните възли, цереброспиналната течност, тъканите на гръбначния и главния мозък.
Географско разпространение. Среща се в екваториална Африка. В огнищата на заболяването трипанозомата е открита в кръвта на антилопи, които служат като негов естествен резервоар. Преносител е африканско кръвосмучещо насекомо - мухата це-це (Glossina palpalis). Заболяването се среща само в района на разпространение на тази муха.
Морфофизиологични особености. Размер от 13 до 39 микрона. Тялото е извито, сплескано в една равнина, стеснено в двата края, снабдено с един флагел и вълнообразна мембрана. Храни се осмотично. Размножаването става безполово, чрез надлъжно делене.
Жизнен цикъл. Трипанозомозата, причинена от T. gambiense, е типично трансмисивно заболяване с естествени огнища. Причинителят на трипанозомозата се развива при смяна на гостоприемника. Първата част от жизнения цикъл на трипанозомата протича в храносмилателния тракт на мухата цеце (Glossina palpalis), извън чийто ареал не се среща трипанозома (африканска сънна болест), втората част - в тялото на гръбначни животни (говеда и дребен добитък, прасета, кучета, някои диви животни), както и хора. Когато муха изсмуче кръвта на болен човек, трипанозомите влизат в стомаха му. Тук те се размножават и преминават през редица етапи. Пълният цикъл на развитие е около 20 дни. Мухите, в тялото на които има трипанозоми от инвазивен стадий, могат да заразят човек при ухапване.
Патогенно действие. Пациент с трипанозомоза има тежки лезии на централната нервна система: наблюдавана мускулна слабост, изтощение, психическа депресия, сънливост. Болестното състояние продължава 7-10 години и при липса на лечение завършва със смърт.
Лабораторна диагностика. За лабораторна диагностика се изследват кръв, пунктати от лимфни възли и цереброспинална течност.
Предотвратяване. Личната профилактика се свежда до приемане на лекарства, които могат да предпазят от инфекция при ухапване от муха цеце. Обществената профилактика се състои в унищожаването на носителя. За тази цел местата за размножаване на мухи - храсти в близост до жилището и по бреговете на водните тела - се третират с инсектициди или се изрязват.
Разред полифлагелати (Polymastigina)

Чревна трихомонада (Trichomonas hominis). Причинява чревна трихомониаза.
Локализация. Дебело черво.
Географско разпространение. навсякъде.
Морфофизиологични характеристики. Тялото има овална форма със заострен израстък в задния край, с едно балоновидно ядро. Дължината на тялото е 5-15 микрона, 4 свободни флагела се простират от предния край, вървейки напред, и един, насочен назад, който е свързан с вълнообразната мембрана. Тялото е пронизано в средата от опорен прът, завършващ със заострен шип в задния край на тялото. Устието на клетката се намира близо до ядрото. В цитоплазмата има храносмилателни вакуоли, които служат за смилане на бактерии и чревно съдържание, погълнати от клетъчната уста. Възможно е и осмотично хранене – течни вещества. Размножаването е безполово, чрез надлъжно делене. Способността за образуване на кисти е оспорвана.
Заразяването става чрез зеленчуци и плодове, мръсни ръце, замърсени с изпражнения, съдържащи Trichomonas, непреварена вода.
Патогенно действиене е доказано, тъй като чревната трихомонада се среща в червата на здрави хора и пациенти с чревни заболявания. Има мнение, че чревната трихомонада не причинява заболяване, а само придружава патологични процеси, причинени от други причини.
Лабораторна диагностика. Микроскопско изследване на петна от изпражнения, откриване на вегетативни форми в изпражненията.
Предотвратяване. Лични - измиване на ръцете преди хранене и след посещение на тоалетната, термична обработка на храната и питейната вода, старателно измиване на сурови зеленчуци и плодове, защита на храната и водата от прах и от мухи, които могат да бъдат механични носители на различни патогени. Обществената профилактика се състои в наблюдение на санитарното състояние на водоизточници, хранителни предприятия и магазини за хранителни стоки, обществени места, борба с мухите и разпространение на хигиенни знания.

Урогенитална трихомонада (Trichomonas vaginalis)- причинител на урогенитална трихомонадоза.
Локализация. Пикочните пътища на мъжете и жените.
Географско разпространение. навсякъде.
Морфофизиологични характеристики. Структурата е много подобна на чревната Trichomonas. Отличителни черти са големите размери на тялото (дължината варира от 7 до 30 микрона). Формата на тялото е най-често крушовидна, има 4 флагела, вълнообразна мембрана и опорна пръчка, завършваща с шип.
Патогенно действие. Предизвиква възпалителни процеси в гениталния тракт, които са продължителни. Смята се, че предразполагащият момент за проява на патогенност е наличието в гениталния тракт на определен вид бактерии.
При жените първоначално се засяга влагалището, но впоследствие заболяването става мултифокално. При остро протичане са характерни обилно течно течение, сърбеж и парене. При мъжете по-голямата част от заболяването протича безсимптомно.
Инфекцията става по полов път, както и при използване на спално бельо, бельо, гъба на пациента. Инфекцията е възможна при преглед от гинеколог чрез замърсени инструменти и ръкавици.
Лабораторна диагностика. Микроскопско изследване на петна от отделянето на пикочните пътища. Диагнозата се поставя въз основа на откриване на вегетативни форми в секретите на пациента.
Предотвратяване. се определя от начина на заразяване.

Ламблия (Lamblia intestinalis). Причинява заболяването лямблиоза. Този флагелат е открит от проф Харковски университет D. F. Lamblem през 1859 г
Локализация. Дванадесетопръстника, за втори път може да проникне в жлъчните пътища.
Географско разпространение. навсякъде.
Лабораторна диагностика. Откриване на кисти във фекалиите или вегетативни форми в съдържанието на дванадесетопръстника по време на сондиране.
Предотвратяване. а) термична обработка на храни и питейна вода; варенето убива кистите на лямблиите; б) цялостно измиване на плодове, зеленчуци и други храни, консумирани сурови; в) съхранение на готвена храна и питейна вода в запечатан съд. Трябва да се има предвид, че кистите на Giardia могат да попаднат върху храната с прах и да бъдат въведени в нея от мухи; г) борба с мухите. Мухите посещават сметища за отпадъчни води, помийни ями, сядат на човешки изпражнения, гниеща храна и, оцветявайки телата си в изпражнения, летят в жилището на човек, където сядат на храна, чинии, човешка кожа; д) измиване на ръцете преди хранене и след ходене до тоалетна.




