Ktoré riasy nemajú bičíkové štádiá. Nižšie rastliny alebo riasy. Samotárske voľne žijúce formy
Monadické vegetatívne bunky a monadické štádiá v životnom cykle (zoospóry a gaméty) rias sú vybavené bičíkmi - dlhými a pomerne hustými výrastkami buniek, zvonka pokrytými plazmalemou. Ich počet, dĺžka, morfológia, miesto uchytenia, povaha pohybu sú u rias dosť rôznorodé, ale v rámci príbuzných skupín sú konštantné.
Bičíky môžu byť pripevnené na prednom konci bunky (apikálne) alebo môžu byť mierne posunuté do strany (subapikálne); ich uchytenie je možné na strane bunky (laterálne) a na ventrálnej strane bunky (ventrálne). Bičíky, ktoré sú rovnaké v morfológii, sa nazývajú izomorfný ak sa líšia - heteromorfný. Isocont- sú to bičíky rovnakej dĺžky, heterokontakt- rôzne dĺžky.
Bičíky majú jediný štrukturálny plán. Je možné rozlíšiť voľnú časť (undulipodia), prechodovú zónu, bazálne teleso (kinetozóm). Rôzne časti bičíka sa líšia počtom a usporiadaním mikrotubulov, ktoré tvoria kostru (obr. 2).
Ryža. Obr. 2. Schéma stavby bičíkov rias (podľa: L.L. Velikanov et al., 1981): 1 - pozdĺžny rez bičíkom; 2, 3 - priečny rez hrotom bičíka; 4 - priečny rez unulipodiou; 5 - prechodová zóna; 6 - rez bázou bičíka - kinetozóm
Undulipodium(v preklade z latinského „vlnová noha“) je schopný vykonávať rytmické pohyby podobné vlnám. Undulipodium je axonéma pokrytá membránou. axonéma pozostáva z deviatich párov mikrotubulov usporiadaných do kruhu a páru mikrotubulov v strede (obr. 2). Bičíky môžu byť hladké alebo pokryté šupinami alebo mastigonémami (chĺpky), zatiaľ čo u dinofytov a kryptofytov sú pokryté šupinami aj chĺpkami. Bičíky primesiofytov, kryptofytov a zelených rias môžu byť pokryté šupinami rôznych tvarov a veľkostí.
prechodová zóna. Funkčne hrá úlohu pri posilňovaní bičíka v mieste jeho výstupu z bunky. V riasach sa rozlišuje niekoľko typov štruktúr prechodovej zóny: priečna platňa (dinofyty), hviezdicovitá štruktúra (zelená), prechodová špirála (heterokont), prechodový valec (primnesiofyty a dinofyty).
Bazálne telo alebo kinetozóm. Táto časť bičíka má štruktúru v podobe dutého valca, ktorého stenu tvorí deväť trojíc mikrotubulov. Funkciou kinetozómu je spojenie bičíka s plazmalemou bunky. Základné telá v mnohých riasach sa môžu zúčastniť jadrového štiepenia a stať sa centrami organizácie mikrotubulov.
Koniec práce -
Táto téma patrí:
Morské riasy
Fisheries University.. Inštitút morskej biológie pomenovaný po v Zhirmunsky Dvor.
Ak potrebujete ďalší materiál k tejto téme, alebo ste nenašli to, čo ste hľadali, odporúčame použiť vyhľadávanie v našej databáze diel:
Čo urobíme s prijatým materiálom:
Ak sa tento materiál ukázal byť pre vás užitočný, môžete si ho uložiť na svoju stránku v sociálnych sieťach:
| pípanie |
Všetky témy v tejto sekcii:
Kryty buniek
Bunkové obaly zabezpečujú odolnosť vnútorného obsahu buniek voči vonkajším vplyvom a dodávajú bunkám určitý tvar. Kryty sú priepustné pre vodu a nízke molekuly rozpustené v nej
Mitochondrie
Mitochondrie sa nachádzajú v bunkách eukaryotických rias. Tvar a štruktúra mitochondrií v bunkách rias sú rozmanitejšie ako v mitochondriách vyšších rastlín. Môžu byť okrúhle.
Pigmenty
Všetky riasy sa dobre vyznačujú súborom fotosyntetických pigmentov. Takéto skupiny v taxonómii rastlín majú postavenie oddelení. Hlavným pigmentom všetkých rias je zelený pigment.
plastidy
Pigmenty v bunkách eukaryotických rias sa ako vo všetkých rastlinách nachádzajú v plastidoch. V riasach sú dva typy plastidov: farebné chloroplasty (chromatofóry) a bezfarebné leukoplasty (ami
Jadro a mitotický aparát
Jadro rias má štruktúru typickú pre eukaryoty. Počet jadier v bunke sa môže meniť od jedného do niekoľkých. Vonku je jadro pokryté membránou pozostávajúcou z dvoch membrán, vonkajšej membrány
Monadický (bičíkový) typ štruktúry talu
Najcharakteristickejším znakom, ktorý určuje tento typ štruktúry, je prítomnosť bičíkov, pomocou ktorých sa monadické organizmy aktívne pohybujú vo vodnom prostredí (obr. 9, A). Pohyblivý š
Rhizopodiálny (améboidný) typ štruktúry
Najvýznamnejšími znakmi štruktúry améboidného typu sú absencia silných bunkových obalov a schopnosť améboidného pohybu pomocou qi dočasne vytvorenej na povrchu bunky.
Palmeloidný (hemimonas) typ štruktúry
Charakteristickým znakom tohto typu štruktúry je kombinácia nehybného životného štýlu rastlín s prítomnosťou bunkových organel charakteristických pre monadické organizmy: kontraktilné vakuoly, stigma, turniket
Typ kokoidnej štruktúry
Tento typ kombinuje jednobunkové a koloniálne riasy, nehybné vo vegetatívnom stave. Bunky kokoidného typu sú pokryté membránou a majú protoplast rastlinného typu (tonoplast bez sokrata).
Trichalický (vláknitý) typ štruktúry
Charakteristickým znakom filamentózneho typu štruktúry je filamentózne usporiadanie nepohyblivých buniek, ktoré vznikajú vegetatívne v dôsledku delenia buniek, ku ktorému dochádza prevažne
Heterotrichálny (multifilárny) typ štruktúry
Multifilamentózny typ vznikol na základe vláknitého typu. Viacvláknový talus pozostáva väčšinou z horizontálnych nití, ktoré sa šíria pozdĺž substrátu, ktoré plnia funkciu pripojenia, a zvislých pozdĺž
Parenchymálny (tkanivový) typ štruktúry
Jeden zo smerov evolúcie multifilamentózneho stielky bol spojený so vznikom parenchymálnych stielok. Schopnosť neobmedzeného rastu a delenia buniek v rôznych smeroch viedla k vzniku
Štruktúra typu sifón
Sifonálny (nebunkový) typ štruktúry je charakterizovaný absenciou talu vo vnútri, ktorý dosahuje relatívne veľké, zvyčajne makroskopické veľkosti a určitý stupeň diferenciácie buniek.
Typ konštrukcie so sifónom
Hlavnou črtou sifónového typu štruktúry je schopnosť vytvárať komplexné stielky z primárneho nebunkového sklzu pozostávajúceho z primárnych viacjadrových segmentov. IN
asexuálna reprodukcia
Nepohlavné rozmnožovanie rias sa uskutočňuje pomocou špecializovaných buniek - spór. Sporulácia je zvyčajne sprevádzaná rozdelením protoplastu na časti a uvoľňovaním štiepnych produktov z
jednoduché delenie
Tento spôsob reprodukcie sa vyskytuje iba v jednobunkových formách rias. Najjednoduchšie delenie sa vyskytuje v bunkách, ktoré majú améboidný typ stavby tela. Rozdelenie améboidných foriem
Fragmentácia
Fragmentácia je vlastná všetkým skupinám mnohobunkových rias a prejavuje sa v rôzne formy: tvorba hormogónie, regenerácia oddelených častí talu, samovoľné odpadávanie konárov, opätovný rast
Rozmnožovanie výhonkami, stolónmi, plodovými púčikmi, uzlinami, akinetmi
V pletivových formách zelených, hnedých a červených rias nadobúda vegetatívne rozmnožovanie svoju úplnú podobu, ktorá sa len málo líši od vegetatívneho rozmnožovania vyšších rastlín. Držanie cesty
sexuálnej reprodukcie
sexuálnej reprodukcie u rias je spojená so sexuálnym procesom, ktorý spočíva v splynutí dvoch buniek, výsledkom čoho je vytvorenie zygoty, z ktorej vyrastie nový jedinec alebo dáva zoospóry.
Zmena jadrových fáz
Počas sexuálneho procesu sa v dôsledku splynutia gamét a ich jadier počet chromozómov v jadre zdvojnásobí. V určitom štádiu vývojového cyklu, počas meiózy, dochádza k zníženiu počtu chromozómov, v dôsledku
Endofyty / endozoity alebo endosymbionty
Endosymbionty alebo intracelulárne symbionty sú riasy, ktoré žijú v tkanivách alebo bunkách iných organizmov (bezstavovcov alebo rias). Tvoria akúsi ekologickú skupinu.
Oddelenie modrozelených rias (cyanobaktérií) – sinice
Názov oddelenia (z gréckeho cyanos - modrý) odráža charakteristickú vlastnosť týchto rias - farbu talu, spojenú s pomerne vysokým obsahom modrého pigmentu fykocyanínu. Cyanogen
Poriadok - Chroococcales
Vyskytujú sa ako jednobunkové „jednoduché“ jedince alebo častejšie tvoria hlienovité kolónie. Keď sa bunky delia v dvoch rovinách, objavia sa jednovrstvové lamelárne kolónie. Rozdelenie v troch n
Oddelenie červených rias – rodophyta
Názov odboru pochádza z Grécke slovo rodón ("radón") - ružová. Farba červených rias je spôsobená odlišnou kombináciou pigmentov. Pochádza zo šedej a fialovej
Rad Bangiaceae–Bangiales
Rod Porphyra má stélku vo forme tenkej lesklej platne s hladkými alebo zloženými okrajmi, ktorá pozostáva z jednej alebo dvoch vrstiev tesne spojených buniek. Základňa dosky zvyčajne ide do
Rad Rhodimeniales - Rhodymeniales
Rod Sparlingia (Rhodimenia) - ploché platne do výšky 45 cm, listovité a klinovité, navrchu rozšírené a dlaňovito členité, od svetloružovej alebo svetlooranžovej až po
Objednajte si Coralline - Corallinales
Rod Corallina - kĺbové vejárovité rozvetvené kríky do výšky 10 cm, rozvetvené, vápenaté, od ružovo-fialovej až po takmer bielu. Rozmnožuje sa nepohlavne a pohlavne. Spo
Gigartine objednávka - Gigartinales
Rod Chondrus - husté kožovité chrupavé kríky do výšky 20 cm, 3-4 krát rozvetvené, svetlo žlté, svetlo ružové, fialovo-tmavo červené. Rastie v spodnej časti litorálu a
Rad Ceramiaceae - Ceramiales
Rod Ceramium - jemné nadýchané kĺbové kríky do výšky 10 cm, dichotomicky alebo striedavo rozvetvené, tmavo žlté s ružovkastým odtieňom. Rozvetvenie dvoch až štyroch rádov, konečné
Oddelenie rozsievok – bacilariophyta
Oddelenie sa nazýva Diatoms (z gréckeho di - dva, tome - rez, pitva), alebo Bacillaria (bacil - palica). Zahŕňa jednobunkové solitérne alebo koloniálne org
Oddelenie heterokontných (mnohobičíkových) rias – heterokontophyta
Zdá sa, že všetky heterokonty majú bičíkový aparát. Bičíky sú 2 a na jednom z nich sú veľmi charakteristické rúrkovité trojčlenné perovité výrastky alebo chĺpky - mastigonémy. Je to hotovosť
Systematika
Fosílne kokolity sú známe z druhohorných ložísk a hojne sa vyskytovali počas veľkej časti jury až po kriedu. Prvosiofyty dosiahli svoju maximálnu diverzitu v neskorej kriede,
Oddelenie kryptofytných rias (cryptomonas) – kryptofyta
Oddelenie je pomenované podľa typového rodu Cryptomonas (z gréckeho kryptos – skrytý, monas – jednotlivec). Zahŕňa jednobunkové, pohyblivé, monadické organizmy. Kryptofytové bunky
A B C D E
Ryža. 53. Vzhľad kryptofytné riasy (podľa: G.A. Belyakova et al., 2006): A - Rhodomonas, B - Chromonas, C - Cryptomonas, D - Hylomonas, E - Goniomonas môžu tvoriť
Oddelenie zelených rias – chlorofytov
Zelené riasy sú najrozsiahlejšie zo všetkých oddelení rias, podľa rôznych odhadov majú 4 až 13 - 20 tisíc druhov. Všetky majú zelenú farbu thalli, čo je spôsobené prevahou chlóru
Objednajte si Ulotrix - Ulotrichales
Rod Ulotrix (obr. 54). Druhy Ulotrix žijú častejšie v sladkej vode, menej často v morských, brakických vodných útvaroch a v pôde. Pripájajú sa k podvodným objektom a vytvárajú jasne zelené kríky.
Bryopsidový rad – Bryopsidales
Väčšina druhov sa nachádza v sladkých a brakických vodách. Niektoré z nich rastú na pôde, skalách, piesku a niekedy aj slaniskách. Rod Briopsis - nitkovité kríky do 6-8 s
Objednajte si Volvox - Volvocales
Rod Chlamydomonas (obr. 57) zahŕňa viac ako 500 druhov jednobunkových rias, ktoré žijú v čerstvých, plytkých, dobre vyhrievaných a znečistených vodných útvaroch: rybníky, kaluže, priekopy atď. Atď
Oddelenie Charophytes (Characeae) - Charophyta
Charofyty – rad sladkovodných zelených rias, ktoré viedli k vyššie rastliny. Ide o formy prevažne s vláknitým talom. Talus je často vertikálny, vypreparovaný a nesie
Oddelenie dinofytov (dinoflagellates) - dinophyta
1. Názov oddelenia pochádza z gréčtiny. dineo - otáčať. Kombinuje prevažne jednobunkové monády, menej často kokoidné, améboidné alebo palmeloidné, niekedy koloniálne
Oddelenie euglenozoa - euglenoidy
Rozdelenie mena podľa typového rodu je Euglena (z gréčtiny eu - dobre vyvinuté, glene - zrenica, oko). Kombinuje jednotlivých monadických alebo améboidných zástupcov. Zriedkavo nájdený v
Slovníček pojmov
Autogamia - pohlavné rozmnožovanie, pri ktorom sa dve sesterské haploidné jadrá spájajú do spoločnej cytoplazmy. Autospóra je štruktúra nepohlavného rozmnožovania, ktorá je
Zdá sa, že všetky heterokonty majú bičíkový aparát. Bičíky sú 2 a na jednom z nich sú veľmi charakteristické rúrkovité trojčlenné perovité výrastky alebo chĺpky - mastigonémy. Práve prítomnosť dvoch nerovnakých bičíkov slúžila ako základ pre pomenovanie tejto obrovskej skupiny vodných organizmov. multiflagelatedriasy, alebo heterocontami. Okrem rozdielov v dĺžke sa bičík líši aj morfologicky: hlavný bičík pozostáva z osi a na nej perovito umiestnených brvitých chĺpkov, bočný bičík je hladký a bičíkovitý. Podobné je aj zloženie pigmentov – chlorofylov A A s a početné karotenoidy, ako aj štruktúra chloroplastov. Chloroplasty sú obklopené štyrmi membránami, pričom dve vonkajšie membrány sú spojené s endoplazmatickým retikulom bunky.
V rámci oddelenia Heterocont (Diverzifikovaných) rias je niekoľko skupín rias zjednotených v triede: Hnedá, Zlatá, Žltozelená, Rafidophytes, Sinura, Feotamnia, Eustigma, niektoré boli predtým považované za samostatné oddelenia.
Trieda Hnedé riasy– Phaeophyceae (Fucophyceae)
názov triedy" hnedé riasy“odráža farbu ich talu (z gréčtiny. Rhaios- hnedý). Farba je spôsobená prítomnosťou veľkého množstva hnedých a žltých pigmentov - okrem chlorofylov A A s A β -karotén v ich chloroplastoch je prebytok xantofylov, najmä fukoxantínu.
Mnohobunkové, prevažne veľké riasy, rozšírené najmä v studených vodách oboch hemisfér. Medzi hnedými riasami nie sú známe ani jednobunkové ani koloniálne formy. Je známych asi 1,8 tisíc druhov.
Bunková štruktúra. Bunka u hnedých druhov rias je eukaryotická: vytvorila organely s vlastnými membránami: jadro, mitochondrie, plastidy atď. Zelené pigmenty - chlorofyly A A s- maskované ďalšími: oranžovými karoténmi a žltohnedými xantofylmi; z posledne menovaných prevláda fukoxantín. V bunkách hnedých rias sa nachádza jeden až veľa plastidov. Na povrchu vonkajšej bunkovej membrány bunky hnedých rias zvyčajne vytvárajú tuhú bunkovú stenu.
Jadro je najčastejšie jedno, ale niekedy je jadier niekoľko. U niektorých predstaviteľov sa so zvyšujúcim sa vekom bunky zvyšuje počet jadier. V jadre je vždy jedno jadierko.
Chloroplasty vegetatívnych buniek hnedých rias majú oko (stigma) a pyrenoidy, ktoré môžu byť jedno alebo viac; niektorým druhom chýbajú pyrenoidy. Ocellus, pozostávajúci z 30–40 lipidových guľôčok, funguje ako šošovka, ktorá sústreďuje svetlo na bičíkový opuch, ktorý je fotoreceptorom. Zoospóry a gaméty hnedých rias zvyčajne nemajú oko. V pyrenoidoch sa ukladajú náhradné látky. Chloroplasty sú najčastejšie malé, diskoidné, parietálne. Ich tvar môže byť hviezdicovitý, stuhovitý alebo lamelový. Tvar chloroplastov v bunkách hnedých rias sa môže meniť, keď bunka starne. Chloroplastový obal sa skladá zo štyroch membrán, pričom dve vonkajšie membrány sú spojené s ER. - endoplazmatické retikulum bunky.
Celulóza, ktorá tvorí bunkovú stenu hnedých rias – niekedy nazývaná algullóza – sa svojimi vlastnosťami líši od celulózy vyšších rastlín. Mimo celulózovej steny je vysoko slizovitá pektínová vrstva, tvorená najmä kyselinou algínovou a jej soľami a zlúčeninami s bielkovinovými látkami. Kyselina algínová tiež sa vyskytuje iba v hnedých riasach.
Všeobecné charakteristiky.
Všetky hnedé riasy sú fototrofy. Produkty fotosyntézy - špecifický sacharid chrysolaminarín,alkohol manitol A lipidy. Lipidy sa hromadia vo forme kvapôčok oleja. Chrysolaminarín sa ukladá mimo chloroplastu: nachádza sa v špeciálnych vakuolách v cytoplazme v blízkosti pyrenoidu. Manitol vykonáva rezervu, ako aj osmoregulačnú funkciu; jeho koncentrácia vo vnútri bunky závisí od zmien slanosti vody.
Mladé bunky hnedých rias obsahujú malé a početné vakuoly, ktoré sa vekom v dôsledku fúzie zväčšujú. V cytoplazme existujú špeciálne formácie - fyzódy, ktoré obsahujú florotanínov. Tieto látky sú známe len u druhov hnedých rias. Funkcia florotanínov môže súvisieť s ochranou riasových stielok pred ich konzumáciou zvieratami, ako sú mäkkýše. Florotaníny hnedých rias pravdepodobne inhibujú usadzovanie epifytických rias a živočíchov na povrchu talu. Predpokladá sa, že florotaníny sa podieľajú na ochrane pred poškodením žiarením a na selektívnej absorpcii z vodné prostredie ióny ťažkých kovov. Bezfarebné florotaníny sa na vzduchu oxidujú za vzniku pigmentu fykofeínu, ktorý dáva sušeným hnedým riasam charakteristickú tmavú farbu.
Štruktúra talu. Medzi druhmi hnedých rias neexistujú jednobunkové a koloniálne formy, všetci jedinci sú mnohobunkové. Forma stielok hnedých rias je najrozmanitejšia. Sú to nite, plazivé alebo vertikálne stojace, jednoradové alebo viacradové; masívne pevné alebo kachľové kôry a kôry; tenké slizničné platničky alebo husté kožovité platničky na stonke, jednoduché alebo členité; bubliny a vrecká; Rúry a šnúry; rôzne rozvetvené tenké kríky alebo mohutné kríky. Pripevnenie tali sa vykonáva pomocou rizoidov alebo chodidiel. Na ich udržanie vo vertikálnej polohe vytvárajú veľké bentické formy hnedých rias vzduchové (plávajúce) bubliny naplnené plynom (obr. 29).
Ryža. 29. Vzhľad hnedých rias: 1 - Fucus; 2 - sacharín; 3 - Alaria; 4 - Sargasum
Thalli zástupcov radov Laminaria a Fucus sú najkomplexnejšie spomedzi hnedých rias. Majú znaky tkanivovej diferenciácie so špecializáciou buniek talu (obr. 30). V ich talí sa dá rozlíšiť štekať pozostávajúce z intenzívne zafarbených buniek a jadro pozostávajúce z bezfarebných buniek, ktoré sa často zhromažďujú vo vláknach. V Laminaria sa v jadre vytvárajú aj sitové rúrky a rúrkové vlákna. Jadro plní nielen transportnú funkciu, ale aj mechanickú, keďže obsahuje závity s hrubými pozdĺžnymi stenami. Medzi kôrou a jadrom mnohých hnedých rias môže byť medzivrstva veľkých bezfarebných buniek.
Výška stélka u predstaviteľov hnedých rias je najčastejšie interkalárna (interkalárna) a apikálna (apikálny), zriedkavo bazálny. Interkalárny rast môže byť difúzny alebo môže existovať zóna rastu. Vo veľkých formách sa interkalárny (interkalárny) meristém alebo aktívne deliaca skupina buniek nachádza v mieste prechodu "stopky" do "listovej" čepele. Veľké riasy majú na povrchu talu aj meristematickú zónu, takzvaný meristoderm (akási obdoba kambia vyšších rastlín).
reprodukcie u hnedých rias, vegetatívnych, asexuálnych a sexuálnych. Gaméty sa tvoria v mnohobunkových gametangiách, zoospórach - in 
Ryža. 30. Vnútorná štruktúra Laminaria(podľa: L.L. Velikanov a kol., 1991): A- pozdĺžny rez stonky; B- prierez stonky; IN- plátok taniera so sori zoosporangie
unilokulárna sporangia.
Vegetatívne rozmnožovanie v mnohých druhoch hnedých rias sa môže uskutočňovať na miestach stélky; v druhoch milý Sphacelaria- plodové vetvičky (plodové púčiky); u druhov rodu Fucus- skupina buniek na chodidle, ktoré sú schopné diferenciácie na nový slez.
K nepohlavnému rozmnožovaniu hnedých rias zvyčajne dochádza pomocou zoospór. Niektoré druhy sa rozmnožujú nepohyblivými tetraspórami alebo monospórami. Spóry nepohlavného rozmnožovania sa tvoria v dôsledku meiózy a následných mitóz v unilokulárna sporangia. Sexuálny proces je izogamia, heterogamia a oogamia. Gamety sa tvoria v mnohostennýtieto gametangie- špeciálne skupiny buniek. V každom „hniezde“ gametangia sa tvorí jedna gaméta.
Na hnedé riasy, sexuálne feromóny- rozpustné látky, ktoré koordinujú činnosť buniek pri rozmnožovaní. Feromóny, ktorých je známych najmenej desať, môžu buď stimulovať otváranie anterídií, alebo priťahovať mužské gaméty k ženským. Tiež sa podieľajú na izolácii druhov. Predpokladá sa, že feromóny sa tvoria vo väčšine druhov hnedých rias, ktoré majú sexuálny proces.
Životný cyklus. Hnedé riasy majú dva hlavné typy životných cyklov. Jeden typ je haplodiplobiont. Sporofyty sú diploidné, t.j. Všetky bunky obsahujú kompletnú sadu chromozómov. V unilokulárnych sporangiách sa tvoria spóry nepohlavného rozmnožovania. Pri tvorbe spór nastáva redukčné delenie – meióza. Takáto redukcia sa nazýva sporická. Bunky (spóry) vytvorené po meióze sú haploidné; obsahujú polovičný počet chromozómov. Haploidné zoospóry a tetraspóry vyklíčia do haploidného gametofytu, na ktorom sa tvoria gaméty vo viacbunkových gametangiách. Diploidná zygota vytvorená po splynutí gamét prerastie do diploidného sporofytu.
Ďalším typom je diplobiont; sporofyty, spóry a gametofyty sú diploidné; redukčné delenie nastáva pri tvorbe gamét. Takáto redukcia sa nazýva gametická. Po splynutí haploidných gamét vzniká diploidná zygota, ktorá vyklíči na diploidný sporofyt. Tento typ je bežný u druhov chalúh (obr. 31).

Ryža. 31. Životný cyklus Sacharíny(podľa: L.L. Velikanov et al., 1991): 1 – sporofyt, 2 – sori zoosporangií, 3 – zoospóry , 4 - gametofyt, 5 - anterídium, 6 - oogónium, 7 - anterozoid, 8 - zygota (oplodnené vajíčko)
Na makroskopickom sporofyte Sacharíny, v hornej časti dosky sa vytvárajú pruhy sori(skupiny) zoosporangií. V zoosporangiách sa po redukčnom delení vytvárajú zoospóry s dvoma nerovnakými bičíkmi. Rastú do mikroskopických gametofytov: samec s anterídiami a samica s oogóniou. Vajíčko oplodnené anterozoidmi okamžite, bez obdobia vegetačného pokoja, vyklíči a vytvorí sporofyt. V tomto životnom cykle Sacharíny končí.
Riasy Fucus nemajú sporofyty. O Fukusa mužské a ženské pohlavné orgány - anterídia so spermiami a plodnice s oogóniou a vajíčkami - sú umiestnené vo výklenkoch talu - scaphids, mužské a ženské scaphidia môžu byť na rovnakom nádoba alebo v rôznych receptoch. Oogonia sedí na nohe klietky , objavujúce sa na stene conceptcula(obr. 32, C, D). Bunka oogónia podlieha trom deleniam

Ryža. 32. Rozmnožovanie Fukusa(podľa: B. Fott, 1971; W. Braune a kol., 1974): A – fragment talu (1 - vzduchová bublina, 2 - recept); B– rez cez nádoby; IN– časť ženskej skafidie; G- oogónia; D- uvoľnenie vajec; E– časť samčieho scaphida; A– vetva s antheridia; 3 – výdaj spermií
jadier, prvým z nich je redukcia, v dôsledku čoho sa v oogóniu vytvorí 8 vajíčok.
Antherídia sa zvyčajne tvoria na rozvetvených vláknach, ktoré ich podporujú - parafýzy(obr. 32, E,A). V dôsledku niekoľkých delení sa vytvorí 64 spermií. spermie Fukusa majú oko umiestnené vo vnútri redukovaného chloroplastu. Na prednom konci spermie sa tvorí 13 mikrotubulov tulákprúd- špeciálna štruktúra, ktorá sa má podieľať na definícii ženského sexuálneho atraktantu.
Spermie sú priťahované k vajíčku Fukusa fukoserraten, nešpecifický feromón vylučovaný vajíčkom. Rozpoznanie na úrovni druhu je spojené so špecifickými oligosacharidmi na vajíčku a spermiách. Na povrchu vajíčka sú rôzne oblasti, z ktorých každá obsahuje špeciálne organické zlúčeniny - glykoproteíny. Spermie obsahujú glykoproteíny usporiadané do miest na membráne a výrastkoch predného bičíka, ako aj na povrchu samotnej spermie. Pri stretnutí s vajíčkom sa spermie pohybuje po škrupine vajíčka, akoby ho skúmala pomocou predného bičíka. Ak sú glykoproteíny spermií komplementárne ku glykoproteínom vajíčka, potom dochádza iba k rozpoznaniu spermie vajíčkom.
Systematika
Fosílne nálezy, ktoré môžu súvisieť s hnedými riasami, sú známe zo stredného silúru (asi 425 ma). Ale tieto nálezy sú tiež podobné niektorým moderným zeleným a červeným riasam. Fosílne nálezy, ktoré možno jednoznačne spájať s modernými hnedými riasami, pochádzajú z miocénu (5–25 miliónov rokov). Tieto riasy pripomína modernú Dictiotu. Vek hnedých rias stanovený molekulárnymi metódami je 155–200 miliónov rokov.
Na rozlíšenie radov hnedých rias sa používa typ organizácie talu, prítomnosť alebo neprítomnosť pyrenoidu, spôsob rastu, typ sexuálneho rozmnožovania (izogamia, heterogamia, oogamia) a životný cyklus. V posledných rokoch bol systém hnedých rias aktívne revidovaný. V rôznych klasifikačných schémach sa ako súčasť tried hnedých rias rozlišuje 7 až 14 rádov.
Desmarestiánsky poriadok–Desmarestiales. Riasy sú huňaté, viacnásobne rozvetvené, s terezovitým, plochým alebo splošteným talom. Pripevňuje sa k podkladu pomocou dobre vyvinutej podrážky. Na povrchu talu sa vyvíjajú chĺpky.
Rod Desmarestia (obr. 33). Hnedé alebo olivové kríky s dobre definovaným hlavným výhonkom, do výšky 0,8–1 m. Prichytené podrážkou v tvare kužeľa. Rozvetvenie 3-4 rádov, striedavé alebo opačné. Vetvy posledných rádov sú ploché alebo zriedkavejšie nitkovité. Druhy sú morské, žijú v arktických a miernych zemepisných šírkach v hĺbke 1–20 m, tvoria húštiny alebo rastú jednotlivo.

Ryža. 33. Desmarestia (r. Desmarestia)
Ralphsovský poriadok–Ralfsiales. Zástupcovia rádu majú talus vo forme kôr odlišná štruktúra alebo pozostávajú z bazálnej kôry a zvislých výhonkov.
Rod Analipus. Thallus sa skladá z trvalej kôrovitej bazálnej časti a z nej vybiehajú jednoročné olivovohnedé zvislé výhonky vysoké až 40 cm.Mnohobunkové výtrusnice sú valcovitého tvaru a dvojradovej štruktúry. Unilokulárne sporangie sú oválne a vyvíjajú sa medzi asimilačnými vetvami. Druhy rodu sú rozšírené v moriach miernych zemepisných šírok a tvoria pásy v prílivovej zóne.
Objednajte si Dictyota–Dictyotales.Pre druhy radu je charakteristický vrcholový rast. Vegetatívne bunky obsahujú početné diskovité chloroplasty bez pyrenoidov. Parenchymálny talus, charakteristické je izomorfné striedanie generácií. Pohlavný proces je oogamný, nepohlavné rozmnožovanie nepohyblivými spórami, spermie majú len jeden bičík. Bičík je dlhý, so sperenými výrastkami a smeruje dopredu. Zástupcovia rádu sa nachádzajú hlavne v tropických a menej často v miernych moriach.
Rod Dictyota (príloha, 4A). Má stélku vo forme plochých olivovohnedých, dichotomicky rozkonárených kríkov vysokých 11–15 cm.Vetvy sú lineárne, úzke, bez žiliek, s vidlicovitými vrcholmi. Tetrasporangia tvoria sori na povrchu talu, ktoré majú neurčitý alebo prstencový tvar. Distribuované v morských vodách miernych a teplých zemepisných šírok.
Objednajte si Laminaria–Laminariales. Laminaria majú stélku pozostávajúcu z dosky, stonky a upevňovacích orgánov - podošvy alebo rizoidov. Platnička je rovnomerná alebo zvrásnená, celá alebo členitá, s dierami, záhybmi, guľami alebo bez nich, od niekoľkých desiatok centimetrov do 20–40 m. Stonka je rozkonárená alebo častejšie nerozvetvená. Sporofyty sú jednoročné alebo viacročné, niektoré druhy majú 11–18 rokov. Slizničné kanály sa nachádzajú v kôre mnohých rias.
Najpopulárnejšími predstaviteľmi hnedých rias sú kelp (druhy rodov Laminaria alebo Saccharina), alebo „morské riasy“, ktoré sa distribuujú najmä v Tichý oceán. Nachádzajú sa od hladiny (okraja) vody do hĺbky 200 m. Laminárie rastú prednostne v hĺbke 2–20 (30) m, na miestach s neustálym pohybom vody, prichytávané na kameňoch a skalách. Laminaria tvoria pásy rias pozdĺž pobrežia morí Ďalekého východu. Najsilnejšie húštiny sa nachádzajú v blízkosti Kurilských ostrovov a približne. Hokkaido, kde je sústredená asi polovica svetových zdrojov laminárií. Ročná svetová úroda chaluhových rias dosahuje 2 milióny ton čerstvej hmotnosti, pričom viac ako 1 milión ton pochádza z produkcie jej marikultúry v Číne.
Rod Sacharín (Laminaria). Thallus je dlhý od 0,5 do 5–10 m, má hrubú lesklú platňu, pružnú stonku a husto prepletené rizoidy, ktoré sú pripevnené k rôznym pevným substrátom. Okraje taniera sú hladké alebo zvlnené, na povrchu taniera sú často vydutia a záhyby. Na oboch stranách platničky sa tvoria nahromadenia sporangií - sori, po uvoľnení spór dochádza k deštrukcii vrchnej časti platničky. Žije od dvoch do niekoľkých rokov. Sacharíny sú cennou priemyselnou surovinou a predmetom marikultúry. Saccharina japonská tvorí húštiny v severnej polovici Japonského mora, ako aj v blízkosti južného a juhovýchodného pobrežia Sachalinu a južných Kurilských ostrovov. Saccharina japonská je najcennejším obchodným druhom, má vynikajúcu chuť a používa sa na prípravu rôznych jedál, konzerv, cukroviniek.
Rad Sphacelariae–Sphacelariales.
Hnedé riasy patriace do tohto rádu majú rozvetvený talus s vrcholovým rastom. Chlpaté konáre. Vegetatívne bunky majú početné chloroplasty bez pyrenoidov. Životný cyklus väčšiny s izomorfnou zmenou generácií, sexuálnym procesom je izogamia alebo heterogamia. Riasy tvoria malé hnedé trsy do výšky 3 cm a rozvetvené kríky do výšky 20 cm na kameňoch alebo iných riasach; existujú endofytickí zástupcovia.
Rod Sphacelaria. Má tenký talus. Vetviace sa vlákna tvoria malé, do 0,5–3 cm vysoké, hnedé trsy. Vegetatívne rozmnožovanie sa uskutočňuje špecializovanými, často trojlúčovými odnožami - propaguly. Sexuálny proces je izogamný. Ženské gaméty vylučujú feromóny ectocarpen a multifiden. Druhy rodu sú distribuované v moriach polárnych aj tropických zemepisných šírok.
Objednajte si Scytosiphon–Scytosiphonales. Zástupcovia s črevným, vezikulárnym, lamelárnym alebo vakovým, často dutým stélom s dĺžkou až 25 cm. Kôra je jednoradová alebo dvojradová, vnútorná časť (stena) talu je tvorená niekoľkými vrstvami veľkých buniek. Na povrchu talu sa vyvíjajú mnohobunkové sporangie, ktoré sa zhromažďujú v sori, ktoré podopierajú ich vlákna - parafýzy a chumáče chlpov.
Rod scytosifón je tenkostenná hnedá rúrka dlhá 4–25 cm (príloha 4B). Talus je pripevnený malou podrážkou a má zriedkavé priečne zúženia. Vyskytuje sa v pobrežnej zóne morí v rôznych zemepisných šírkach, často tvorí pásy v prílivovej zóne.
Fucus poriadok–Fucales. Majú trvalú stélku vo forme veľkých nepravidelne alebo dichotomicky sa rozvetvených kríkov do dĺžky 1–3 m. K podkladu sú pripevnené podrážkou v tvare kužeľa. Thallus má výhonky, fyloidy (listovité útvary), schránky (zásobníky gamét) a plávacie (vzduchové) mechúre. Nachádzajú sa v prílivovej zóne všetkých miernych a arktických morí, kde tvoria silné húštiny - pásy.
Rod sargasum(obr. 34, IN). Bohato rozvetvené olivovohnedé alebo hnedé kríky vysoké 0,5–1,5 m. Prichytené kužeľovou podrážkou. Fyloidy sa vyvíjajú na bočných výhonkoch vybiehajúcich z valcovej stonky v dolnej a strednej časti talu. Fyloidy sú kožovité, so stredovým rebrom, 0,2–2 cm dlhé. Plavecké mechúre sú oválne alebo zaoblené, s výrastkami alebo bez nich. Sargasy sú rozšírené v miernych a subtropických zemepisných šírkach od spodného litorálu do hĺbky 10 m, kde vytvárajú husté alebo riedke húštiny. V južnom Tichom oceáne – takzvanom Sargasovom mori – tvoria rozsiahle plávajúce akumulácie, v ktorých sa rozmnožujú len vegetatívne.
Rod Fucus (obr. 32).Thallus kožovitá, huňatá, konáruje sa dichotomicky, 25–50 cm vysoká, pri pobreží Ďalekého východu rastie niekoľko rokov, žije v prímorskej zóne. V plytkej vode často vytvára veľké húštiny. Druhy tohto rodu sa vyznačujú dichotomickými
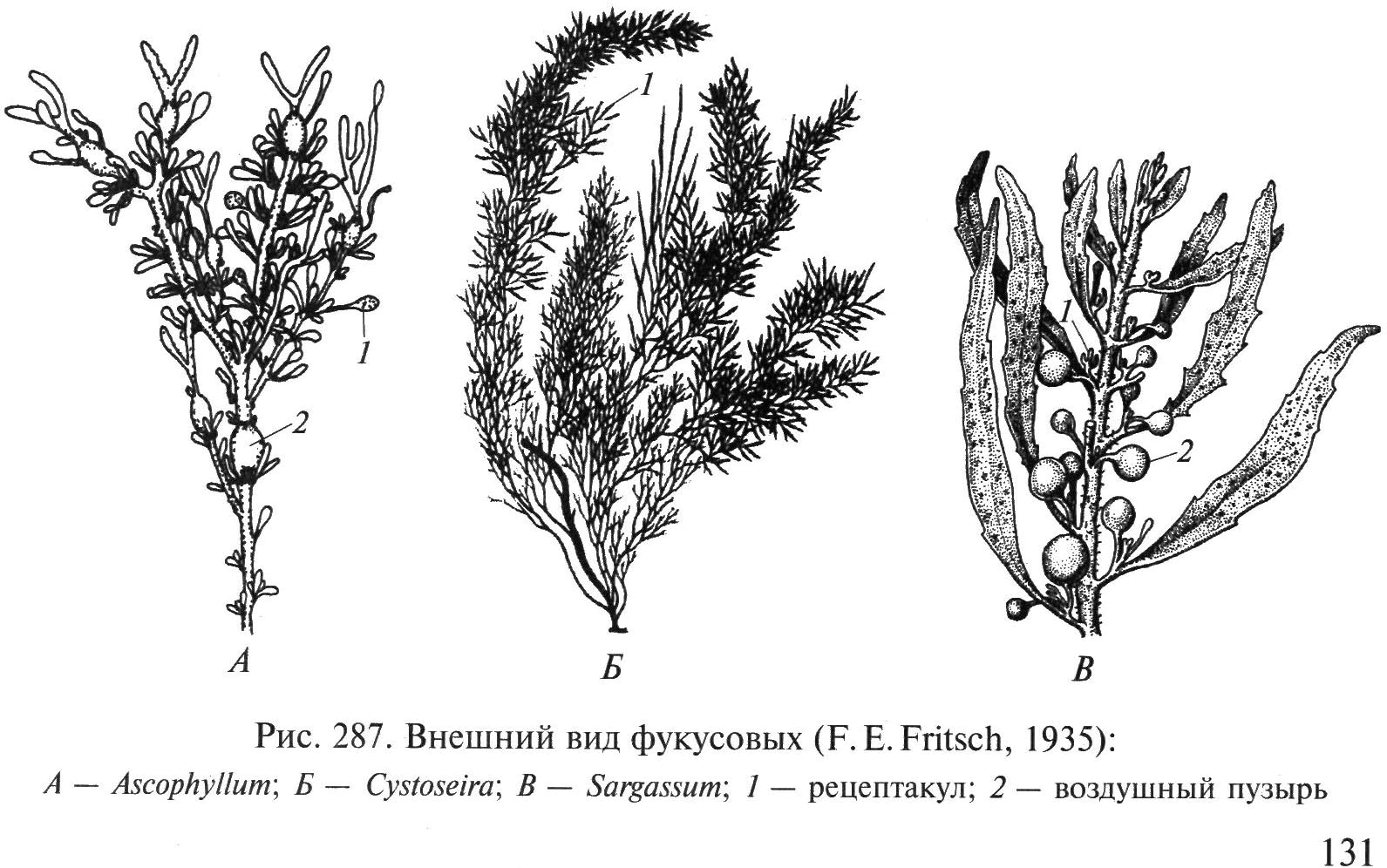
Ryža. 34. Vzhľad riasy Fucus: A – Ascophyllum; B – Cystoseira; IN – sargasum; 1 - nádoba, 2 - plavecký mechúr
rozvetvený talus s plochými vetvami s pozdĺžnym rebrom, ktorý je pripevnený ku kameňom kužeľovou podrážkou. Fukusy sa používajú ako hnojivá, ako krmivo pre hospodárske zvieratá, na výrobu kŕmnej múky, alginátov, používajú sa v medicíne na liečbu strumy, korekciu hmotnosti atď.
Objednajte si ektokarpy–Ectocarpales. Riasy, ktoré sú tu zahrnuté, sú mikroskopické, voľne žijúce, epifyty, epizoity alebo endofyty. Reprodukčné orgány sú zastúpené unilokulárnymi (sporangia) a multilokulárnymi (gametangia) sporangiami; sú dlhovalcovité, krátkovalcovité alebo strukovité.
Rod ektokarpus. Thallus vo forme mäkkých hnedých kríkov do výšky 2 cm. Kríky sú tvorené jednoradovými, striedavo, dichotomicky alebo nesprávne rozvetvenými vláknami. Mnohobunkové výtrusnice sú strukovité, vytvorené na bočných konároch na 1–3-bunkovej stopke. Druhy sú rozšírené v moriach rôznych zemepisných šírok, v čistých a znečistených vodách, v litoráli a v hornej časti sublitorálu. Epifyty sa nachádzajú aj v znečistení antropogénnych substrátov.
Ekológia
Prevažná väčšina hnedých rias žije v morských vodách. V sladkých vodách sa vyskytuje iba 8 druhov. Existujú ročné aj viacročné druhy, ktorých vek môže dosiahnuť 15–18 rokov. Laminaria riasy Nereocystis A Macrocystis, ktoré žijú pozdĺž tichomorského pobrežia Severnej Ameriky, majú maximálnu veľkosť pre všetky riasy - až 30–40 m na dĺžku. Tvoria obrovské podmorské lesy v moriach Ďalekého východu. Hnedé riasy sa viažu na širokú škálu substrátov - kamene, kamene, štrk, lastúry a lastúry morských živočíchov a iné riasy. Niektoré malé formy hnedých rias žijú vo vnútri tkanív iných rias ako endofyty. Väčšina hnedých druhov žije v pripojenom stave. Tále odtrhnuté od zeme sú prúdom unášané do pokojných miest s bahnitým dnom a tam naďalej existujú. Druhy so vzduchovými bublinami na taluse, ktoré sa odtrhávajú od zeme, plávajú na povrch a vytvárajú veľké plávajúce zhluky, najmä v oblastiach so stabilným kruhovým prúdom, ako napríklad v Sargasovom mori. V takýchto zhlukoch existujú pomerne dlho a rozmnožujú sa len vegetatívne.
Hnedé riasy sú rozšírené vo všetkých moriach, no najväčší rozvoj dosahujú v moriach miernych a subpolárnych šírok. Ich veľké stélky rastú pripevnené na skalách a kameňoch a na tichých miestach blízko pobrežia a vo veľkých hĺbkach sa môžu dokonca držať na ventiloch lastúr mäkkýšov a štrku. Môžete ich stretnúť v rôznych hĺbkach – od prímorskej zóny, kde sú pri odlive celé hodiny mimo vody, až po 40–200 m. V oblasti Havajských ostrovov v hĺbke 180 m druhy rodu sargasum, A Kelp V Jadranské more bolo nájdené v hĺbke 200 m. Najhojnejšie húštiny hnedých rias sú pozorované v hĺbke 6–15 m, kde sú najlepšie svetelné podmienky a neustály pohyb vody, ktorý prináša živiny do ich slín a limitov usídlenie bylinožravých zvierat - fytofágy.
Trieda Zlaté riasy -Chrysophyceae
Jednobunkové, koloniálne alebo mnohobunkové organizmy, ktoré majú vo svojom životnom cykle zvyčajne bičíkové štádium. Bunky sú jednojadrové, s jedným alebo viacerými chloroplastmi. Zvyčajne je prítomná stigma (obr. 36). Bičíky 1–2, nerovnaké. Kontraktilná vakuola sa nachádza v prednej časti bunky. Vonkajšie obaly buniek týchto organizmov sú veľmi rôznorodé. U najprimitívnejších druhov je bunka odetá do jemného periplastu, ktorý umožňuje tvorbu výbežkov rôznych tvarov (rizopódia, pseudopódia), u iných je plazmalema pokrytá tvrdou celulózovou membránou, niekedy silne slizovou. Domy v zlatých riasach majú rôzne tvary: vázovité, guľovité, vajcovité, valcové; s jedným alebo viacerými otvormi. Klietka je pripevnená k základni domčeka ohybnou nohou alebo môže byť voľná. U niektorých chryzofytov sa na povrchu bunky ukladajú vápnité útvary – kokolity – rôznych tvarov a veľkostí a kremíkové bičíkovce majú vnútornú kremíkovú kostru. Rozmanitosť štruktúry týchto kostier umožňuje využiť pozostatky rôznych foriem fosílnych silikoflagelátov na určenie veku geologických hornín, ktoré ich obsahujú. U rozvinutejších predstaviteľov oddelenia zlatých rias sú bunky oblečené v škrupine pozostávajúcej z kremíkových šupín (niekedy nesúcich hroty) alebo sú uzavreté v domoch, cez otvory ktorých vychádzajú bičíky alebo pseudopódia.
Domy v druhoch zlatých rias sú vázovité, guľovité, vajcovité, valcovité, s jedným alebo viacerými otvormi. Klietka je pripevnená k základni domčeka ohybnou nohou alebo môže byť voľná.
Pigmenty - chlorofyly A A s fukoxantín. Chryzofyty sa vyznačujú najvyšším obsahom fukoxantínu spomedzi heterokontov (55 – 93 % všetkých karotenoidov).
Väčšina druhov sú sladkovodné fototrofy, ale nachádzajú sa aj heterotrofy a fagotrofy.
Náhradné látky - chrysolaminarín a oleje.
Medzi jednobunkovými formami zlatých rias prevládajú sladkovodné planktonické druhy a medzi mnohobunkovými formami bentické alebo epifytické. Nachádzajú sa hlavne v chladných morských a sladkých vodách. Je známych asi 360 druhov.

Ryža. 36. Zlaté riasy (podľa: C. Hoek van den et al., 1995): A- bunková štruktúra; B- radikulárny systém; IN- tripartitný mastigoném; G- rez časti chloroplastu; 1 - bazálne telo; 2 - bazálny opuch; 3 - krátky bičík; 4 - stigma; 5 - rizoplast; 6 - jadro; 7 - jadrová membrána; 8 - jadierko; 9 - mitochondrie; 10 - vakuoly; 11 - hlien; 12 - plazmalema; 13 - vezikuly; 14 - slizničné telo; 15 - lipidy; 16 - chloroplast; 17 - Golgiho prístroje; 18 - kontraktilná vakuola; 19 - dlhý bičík: 20 - 1. koreň; 21 - 2. chrbtica; 22 - 3. chrbtica; 23 - 4. koreň; 24 - chloroplastový nukleoid; 25 - HES; 26 - chloroplastová škrupina; 27 - pásová lamela; 28 - lamela; 29 - dlhé bočné vlákno; 30 - krátke bočné vlákno; 31 - koncové vlákno; 32 - tubulárna časť mastigonému
štruktúra talu u zlatých rias je najčastejšie monadická, ale môže byť veľmi rôznorodá: kokoidná, palmeloidná, plazmoidná, améboidná, filamentózna, parenchýmová (obr. 37).

Ryža. 37. Zlaté riasy (podľa: L.L. Velikanov et al., 1981): A – Ochromonas: 1 - vzhľad, 2 - cysta; B – Chromulina: 1 - vzhľad, 2 - cysta, 3 - schéma tvorby filmu z cýst; IN – Dinobrion: 1 - celkový pohľad na kolóniu, 2 - cysta; G – Chryzameba; D – gibberdia; E – Hydrurus: 1 - celkový pohľad na kolóniu, 2 - hrot vetvy, 3 - zoospóra
reprodukcie. V zlatých riasach existujú tri spôsoby reprodukcie: vegetatívna, asexuálna a sexuálna.
Vegetatívne rozmnožovanie sa vyskytuje pozdĺžnym delením buniek alebo rozpadom kolónie na časti v dôsledku delenia buniek v jednom, dvoch alebo troch smeroch.
sexuálny proces(bunky zlatých rias sú diploidné) - izogamia alebo konjugácia. V dôsledku sexuálneho procesu, ako aj za nepriaznivých podmienok, sa vytvárajú cysty s hustou škrupinou obsahujúcou kremík.
asexuálna reprodukcia realizované jedno- alebo dvojbičíkovými zoospórami, ktoré sa vyvíjajú v bežných vegetatívnych bunkách alebo zoosporangiách. Sexuálny proces: holo-, izo- a autogamia. Hologamia je typ sexuálneho procesu, pri ktorom sa dve bunky podobné vegetatívnym bunkám zlúčia na predných koncoch do dvojjadrovej zygoty, ktorá sa potom zmení na silicizovanú cystu.
Systematika
Zo zlatých rias bola koncom minulého storočia na základe výsledkov molekulárno-genetických štúdií izolovaná trieda Sinuraceae. Rozlišuje sa aj trieda Feotamniaceae - od žltozelených a zlatých rias. Preto sa rozsah triedy Chrysophyceae zmenšil av súčasnosti zahŕňa asi 360 druhov zaradených do troch rádov: Chromulinales, Hibberdiales a Hydrurales. Tvar, štruktúra a počet bičíkov, ako aj pigmentové zloženie a stavba tela sú dôležité systematické znaky.
Chromulínový poriadok - Chromulinales. Tento rád zahŕňa organizmy s monadickým, palmeloidným a améboidným typom diferenciácie talu. Monadické bunky s jedným bičíkom viditeľné pod svetelným mikroskopom.
Rod Chryzameba(obr. 37, A) - sladkovodné améboidné riasy, vo svojom životnom cykle majú bičíkové štádium. Zygota sa vyvinie do stomatocysty.
Rod Chromulina(obr. 37, IN) - jednobunkové voľne plávajúce organizmy, ktoré žijú hlavne v sladkých vodách. Bunky sú nahé; existuje jeden alebo dva zlaté plastidy.
Rod Dinobrion(Obr. 37 , G)- jednobunkoví a koloniálni, voľne plávajúci alebo pripútaní zástupcovia. Monády sú umiestnené vo vázovitých domčekoch tvorených celulózovými mikrofibrilami. Okrem celulózy obsahuje domček aj veľké množstvo aminokyselín. Huňatá forma kolónií je spojená s metódou vegetatívneho bunkového delenia, keď sa jedna z dcérskych buniek, opúšťajúca rodičovský dom, prichytí k jej otvoru. Otáčajúc sa okolo svojej osi tvorí svoj vlastný dom. Obe detské bunky môžu opustiť aj rodičovský dom. Bunky obsahujú 1-2 chloroplasty, na prednom konci bunky je viditeľné oko a 2 kontraktilné vakuoly. Chrysolaminarín sa nachádza vo vakuole na zadnom konci bunky. Reprodukcia je vegetatívna a sexuálna.
Objednajte si Gibberdiae–Hibberdiales. Zástupcovia tohto rádu sa vyznačujú jedinečným pigmentovým zložením. Okrem fukoxantínu majú dodatočný karotenoidový pigment, ktorý zachytáva svetlo, antheraxantín.
Zahŕňa pôrod Gibberdia, Chromophyton, Styloceras, Chrysopixis, Platyteka a ďalšie.
Rod gibberdia má dve štádiá životného cyklu: koloniálnu palmeloidnú nehybnú a jednobunkovú monadickú pohyblivú. Pri svetelnej mikroskopii je v monadickom štádiu viditeľný iba jeden bičík. Okrem fukoxantínu je charakteristická prítomnosť ďalšieho doplnkového svetlozberného karotenoidového pigmentu, antheraxantínu.
Objednajte si Hydrorus–Hydrurales. Zástupcovia tohto rádu majú talus s palmeloidnými a pseudoparenchymálnymi typmi štruktúry. Charakteristická je prítomnosť jedinečných tetraedrických zoospór. Nie je tam oko. Mitóza je polouzavretá.
Zahŕňa pôrod Hydrorus, Celloniella, Federmatium, Chrysonebula a ďalšie.
Rod Hydrurus(obr. 37, E) je medzi palmeloidnými riasami najviac diferencovaná. Jeho stélka sú veľké, až 30 cm dlhé slizké kolónie, ktoré vyzerajú ako hnedé rozvetvené šnúry, často vydávajú veľmi nepríjemný zápach. V kolóniách je možné rozlišovať medzi hlavným kmeňom a bočnými vetvami. Bunky ponorené do bežného hlienu obsahujú jeden miskovitý chromatofór a niekoľko kontraktilných vakuol. Bunky sú voľne umiestnené pozdĺž okraja kolónie a hustejšie v strede. Talus môže rásť len delením apikálnych buniek. Pri nepohlavnom rozmnožovaní sa v bunkách bočných vetiev kolónie tvoria zoospóry. Vytvára sférické cysty. Vyskytuje sa v horských potokoch a riekach so studenou vodou, kde sa prichytáva na tvrdých substrátoch.
Trieda rias Sinura -Sinurophyceae
syn- spolu a ura- chvost. Táto skupina organizmov bola izolovaná v roku 1987 zo zlatých rias. Zjednocuje monadické jednotlivé a koloniálne organizmy, niekedy so striedaním v životnom cykle monadického a palmeloidného štádia. Povrch pektínových škrupín je pokrytý škrupinou z kremíkových vločiek. Mitochondrie s tubulárnymi kristami sa zvyčajne nachádzajú v cytoplazme okolo chloroplastu. Jadro je jedno, chloroplasty obklopené štyrmi membránami, zvyčajne dvoma. Lamely sú tritylakoidné, existuje pásová lamela. Hlavnými pigmentmi sú chlorofyly A A s, β -karotén a fukoxantín. Neexistuje žiadna stigma. Bunky zvyčajne s 2 nerovnakými bičíkmi. Dlhý perovitý bičík smeruje dopredu. Krátky, hladký bičík, niekedy silne redukovaný, smerovaný dozadu.
Bunky sa rozmnožujú hlavne pozdĺžnym delením. Kolónie, ktoré sa rozpadajú, vedú k vzniku mladých kolónií. U niektorých druhov je sexuálny proces opísaný vo forme izogamie. Zároveň, na rozdiel od zlatých rias, k fúzii izogamét v sinurických riasach nedochádza na predných, ale na ich zadných koncoch. Silicifikované cysty s pórmi sa tvoria endogénne, podobne ako u chryzofytov.
Súčasťou bentosu sú prevažne planktónne formy, palmeloidné štádiá. Väčšina druhov rias sinura sú fototrofné sladkovodné organizmy.
Systematika.
Trieda rias Sinura zahŕňa 7 rádov: Chloramoebales, Synurales, Rhizochloridales, Ochromonadales, Heterogloeales, Parmales a Thallochrysidales.
Objednajte si Sinura - Synurales. Monadické formy, pektínové bunkové membrány sú zvyčajne pokryté plášťom kremičitých vločiek, stmelených organickou hmotou do jedného puzdra. Chloroplasty jeden alebo dva.
Rod Sinura(Obr. 38) - sladkovodné monadické formy s dvoma nerovnakými bičíkmi a dvoma parietálnymi chloroplastmi. Jadro je hruškovitého tvaru, nachádza sa v prednej časti bunky. Za jadrom je jedna veľká vakuola a v zadnej časti bunky je niekoľko malých kontraktilných vakuol. Bunky sú pokryté šupinami, ako dlaždice.Rozmnožovanie je najčastejšie vegetatívne, u niektorých je známy sexuálny proces. Vytvára kolónie (obr. 39, A). Často sa cysty objavujú vo všetkých bunkách kolónie. S masovým rozvojom Sinura môže spôsobiť nepríjemný zápach vody.

Ryža. 38. Vzhľad buniek Sinura
Rad Rhizochlorid - Rhizochloridales. Prevažujú sladkovodné organizmy s rhizopodiálnym typom diferenciácie talu, žijúce hlavne v sladkovodných útvaroch.
Rod Mallomonas(obr. 39, B-D) – jednobunková monáda s jedným dobre vyznačeným bičíkom a jedným rozdvojeným chloroplastom.

Ryža. 39. Vzhľad sinurických rias (podľa: G.A. Belyakova et al., 2006): A – štiepna kolónia Sinura; B, C – tvorba statospór a G – vzhľad Mallomonas
Bunka je pokrytá kachľovými a špirálovitými šupinami, niektoré majú ihlovité ostne (obr. 39, G). Charakteristická je prítomnosť statospór (obr. 39, B, C). U mnohých predstaviteľov je opísaný sexuálny proces, hologamia.
Rod Mixochlorisžije v bunkách listov sphagnum machov, má vzhľad veľkého mnohojadrového plazmódia. Na jeseň tvorí cysty, ktoré klíčia na jar. Vychádzajú z nich zoospóry alebo améby, ktoré prenikajú do prázdnych buniek listov a tam sa spájajú do plazmódia.
Rad Chloramoebaceae – Chloramoebales. Zahŕňa zástupcov monád. Nachádza sa v slaných a sladkých vodách.
Rod heterochloris – jeho bunky môžu meniť tvar a vytvárať pseudopódia. Tento jav je vlastný mnohým predstaviteľom rádu, ako aj tendencii k živočíšnemu spôsobu výživy. V bunke je niekoľko chloroplastov, olejových kvapiek a chrysolaminarínu. Pred bunkou sú kontraktilné vakuoly, v strede - jedno jadro. Rozmnožuje sa vegetatívne delením buniek.
Objednať Ochromonadales - Ochromonadales. Kombinuje holé formy s dvoma nerovnakými bičíkmi. Sladkovodné a morské formy.
Rod Ochromonas zahŕňa jednobunkové monády s dvoma nerovnakými bičíkmi. Bunky sú obalené iba plazmalemou. Na prednom konci bunky je kontraktilná vakuola a oko, na zadnom konci je vakuola s chrysolaminarínom. Rozmnožuje sa vegetatívne, štiepna brázda začína na prednom konci bunky medzi dvoma pármi bičíkových báz. Druhy rodu sú bežné v oligotrofných sladkých vodách, ale existujú aj morskí zástupcovia.
Rad Heterogleyes - Heterogloeales. Zahŕňa riasy s palmeloidným typom diferenciácie talu. Zástupcovia rádu sa vyskytujú častejšie v sladkých ako v slaných vodách.
Rod Helmintogleažije v brakických vodách. Táto kolónia sediaca na rozšírenej základni pozostáva z rozvetvených slizkých pásov. V tomto hliene sú náhodne usporiadané protoplasty, z ktorých každý je obklopený vlastným hlienovým obalom.
Trieda rias Phaeotamnia - Phaeothamniophyceae
Názov triedy pochádza z rodu typu Phaeothamnion(z gréčtiny. phaeos- hnedý ker). Zástupcovia tejto skupiny organizmov boli identifikovaní v oddelenej triede od žltozelených a zlatých rias v roku 1998 na základe analýzy sekvencie ribozomálnych génov a znakov bunkovej ultraštruktúry. Riasy Feotamnia sa vyznačujú unikátnou kombináciou pigmentov: fukoxantín s heteroxantínom, absencia violaxantínu.
Vlastnosti bunkovej štruktúry: absencia vakuol s chrysolaminarínom a absencia endogénnych cýst so stenami oxidu kremičitého.
Trieda zahŕňa jednobunkové, koloniálne a mnohobunkové organizmy s kokoidným, palmeloidným a filamentóznym typom štruktúry talu. Bičíkovité štádiá s dvoma nerovnakými bičíkmi. Bičíky laterálne alebo subapikálne. Stigma sa nachádza v zoospórach. Pigmenty - chlorofyly A A s, β -karotén, fukoxantín, diadinoxantín, diatoxantín a heteroxantín. Hlavným náhradným produktom je paramylon ( β -glukán). Bunková stena je prítomná, pri delení buniek vzniká dcérska stena vnútri materskej steny.
V feotamniaceae, z hlavných spôsobov reprodukcie, sa nachádzajú iba vegetatívne a asexuálne. Nepohlavné rozmnožovanie sa uskutočňuje pomocou autospór alebo zoospór. Sexuálna reprodukcia nie je známa.
Druhy rias Feotamnia žijú výlučne v sladkých vodách. Systematika.
V súčasnosti je potvrdená monofyletická trieda rias feotamnia a medzi ochrofytmi má bližšie k hnedým a žltozeleným riasam ako k zlatým riasam. V súčasnosti je známych asi 30 druhov týchto rias, pripisovaných jednej rad Theotamnia - Phaeothamniales.
Rod Theotamnion predstavuje pripevnené vetviace nite do výšky 1 cm (obr. 40). Bunky sú valcovité, nepravidelné, rozšírené nahor, pozdĺž

Ryža. 40. Vzhľad Theotamnion.
dva až trikrát dlhšie ako široké. Počas delenia buniek sa materiál rodičovskej steny spotrebuje na vytvorenie vrstveného obalu obklopujúceho bunky vlákna. Na periférii bunky pod plazmatickou membránou sú vezikuly. Pripomínajú fyzódy – útvary nachádzajúce sa v bunkách hnedých rias. Bunky obsahujú jeden až niekoľko diskovitých olivovo-hnedých plastidov. Zoospóry sú tvorené 1–2, menej často 4–8 v jednej bunke. V zoospórach sú bičíky pripevnené bočne. Poloha bičíkových koreňov Theotamnion pripomína žltozelené a hnedé riasy. Chýba mu tiež rizoplast. V životnom cykle sa môžu vytvárať cysty, ale v ich stenách nie je kremík. Cysty klíčia ako zoospóry. Usadzuje sa ako epifyt na vláknitých riasach. Obýva stojaté a pomaly tečúce sladkovodné útvary.
Trieda Rafid rias -Raphidophyceae
Názov triedy pochádza z gréčtiny rhaphid - ihla. Zjednocuje jednobunkové biflagelátové organizmy bez bunkovej steny. Väčšinou sladkovodné organizmy so sploštenými bunkami (obr. 41).
Bunky majú bičíky nerovnakej dĺžky; bičík smerujúci dopredu je perovitý a dlhší; bičík smerujúci dozadu je hladký a kratší. Prechodná špirála chýba. Chloroplasty sú malé, početné alebo zriedkavejšie 1-2 na bunku, obklopené štyrmi membránami, z ktorých dve sú CES membrány. Tylakoidy sú usporiadané v hromadách po troch. Jadro je veľké, obklopené prstencom diktyozómov. Mitochondrie s tubulárnymi kristami. Existujú kontraktilné vakuoly. Cytoplazma je často vakuolizovaná. Bunky sú holé, obklopené iba plazmalemou. Stigma (oko) zvyčajne chýba.
Fotosyntetické pigmenty - chlorofyly A A s, β -karotén, vosheriaxantín, dinoxantín, diadinoxantín, heteroxantín.
Náhradnými asimilačnými produktmi sú tuky a oleje, menej často škrob a chrysolaminarín.
Žijú tiež v rašelinísk a iných biotopoch s kyslými a neutrálnymi reakciami vody. Niektoré druhy sa nachádzajú v brakických a morských vodách.

Ryža. 41. Vzhľad raphidových rias.
Systematika.
Veľmi dlho bola táto skupina rias vďaka svojej zelenej farbe, prítomnosti hltana a množstvu ďalších štruktúrnych znakov považovaná za triedu Chloromonas medzi euglenoidmi. Údaje z cytológie, biochémie, fyziológie a molekulárnej biológie však ukázali ich nepochybnú príslušnosť k oddeleniu rias Heterocont (Ochrophyte) a monofyletickú povahu skupiny rias Rafid.
Z jedinej je známych asi 25 druhov rad Huttonellaceae– Chattonellales.
Rod heterosigma(obr. 42, A) zahŕňa morské bičíkovce. Bunky oválneho tvaru s bičíkmi posunutými do bočná strana, plastidy sa nachádzajú na periférii bunky. Neexistujú žiadne trichocysty. Plávajú a robia rotačné pohyby okolo pozdĺžnej osi tela. Môže spôsobiť toxické „kvety vody“ v pobrežných vodách.

Ryža. 42. Rafidové riasy (od: R. E. Lee, 1999): A – heterosigma; B – Goniostomum; 1 - chloroplast; 2 - mukocysta; 3 - kontraktilná vakuola; 4 - trichocysta; 5 - jadro
Rod Goniostomum(obr. 42, B) sú mobilné dorzoventrálne stlačené monády. Dorzálna strana je zakrivená, ventrálna strana je sploštená. Brázda na prednom konci vedie do trojuholníkového hltana, z ktorého vychádzajú dva dlhé bičíky; ich dĺžka je porovnateľná s dĺžkou bunky. Plastidy sa nachádzajú na okraji bunky. Pod plazmalemou sú trichocysty. Žije v sladkých vodách s kyslým pH.
Trieda rias Eustigma -Eustigmatophyceae
Názov triedy pochádza z gréčtiny EÚ- "dobre a stigma- "značka", "škvrna". Trieda spája nahé jednobunkové, menej často koloniálne organizmy prevažne kokoidnej štruktúry.
Jedno alebo viac jadier. Chloroplast je zvyčajne jeden, obklopený 4 membránami. Bičík je zvyčajne jedno predné perovité a druhé bazálne telo alebo zriedkavejšie dva nerovnaké bičíky. Stigma (oko) je prítomná, nachádza sa mimo chloroplastu, jeho granuly sú bez membrán.

Ryža. 43. Vzhľad rias Eustigma
Chloroplast je zvyčajne jednoduchý, veľký, viaclaločný, miskovitý alebo parietálny, žltozelenej farby. Tylakoidy sú zoskupené do lamiel v skupinách po troch, obopínajúca lamela chýba.
Hlavnými pigmentmi sú chlorofyl a, β-karotén, violaxantín a vosheriaxantín. Pyrenoid je zvyčajne prítomný iba vo vegetatívnych formách.
Náhradné látky sú oleje a látka neznámeho charakteru, ktorá sa ako pevný materiál ukladá mimo plastidov. Netvorí sa škrob.
Žijú v sladkých vodách, menej časté sú v moriach a v pôde.
Systematika.
Predtým bola táto skupina organizmov klasifikovaná ako žltozelené riasy. V súčasnosti sú riasy Eustigma považované za triedu v oddelení Heterocontophytes. Je známych asi 35 druhov, väčšinou sladkovodných, ktoré sa pripisujú jedinému Rad Eustigma - Eustigmatales.
Rod Eustigmatos(obr. 43) - jednobunkové drobné guľovité riasy. Bunková stena je hladká, celistvá, bez ornamentov. Žltozelený chloroplast jednoduchý, laločnatý, parietálny, so stopkovitými polyedrickými pyrenoidmi. Veľká centrálna vakuola s červeným obsahom. Rozmnožuje sa 2 alebo 4 autospórami alebo fľaškovitými zoospórami s jedným prichádzajúcim bičíkom, veľkou stigmou umiestnenou mimo chloroplastu na prednom konci a jedným zadným chloroplastom bez pyrenoidu. Bežná zložka pôdnej flóry. Známe z Nového Zélandu, Rakúska, Islandu az oblasti Arizony ako „horčičné morské riasy“.
Trieda Žltozelené riasy -Xanthophyceae
Žltozelené riasy sú riasy, ktorých chloroplasty sú zafarbené na svetlo alebo do tmy žltá, veľmi zriedkavo zelená a len občas modrá. Farba stielky je určená prítomnosťou nasledujúcich pigmentov v bunkách chloroplastov - chlorofylov A A s, β -karotén a xantofyly. Prevaha posledného určuje originalitu farby žltozelených rias. Okrem toho paramylon, kvapká olej a len u niektorých druhov sa navyše v bunkách hromadia hrudky leukozínu a volutínu ako hlavný produkt asimilácie. Žltozelené riasy netvoria škrob. Charakteristickým znakom žltozelenej je prítomnosť monadickej štruktúry vo vegetatívnych bunkách a dvoch nerovnakých bičíkov v zoospórach. Bunková stena obsahuje celulózu, glukózu a urónové kyseliny. Bunková stena sa často skladá z dvoch častí.
Rozmnožovanie je vegetatívne, nepohlavné a sexuálne.
Široko distribuovaný v sladkých vodách. Zriedkavo sa vyskytuje v morských, brakických vodách a v pôde.
Predtým sa trieda žltozelených rias nazývala riasy Tribophytium, podľa typu rodu Tribonema (z gréčtiny. tribon – zručný, prefíkaný a nema – vlákno). Je známych asi 450 druhov.
Žlté zelené sa vyznačujú výraznou morfologickou rozmanitosťou. Medzi početnými predstaviteľmi tohto oddelenia sa nachádzajú takmer všetky hlavné typy stavby tela: améboidná, monadická, palmeloidná, kokoidná, vláknitá, multifilamentózna, lamelárna a sifonálna (obr. 44 – 46). Thallus jednobunkový,

Ryža. 44. Vzhľad žltozelených rias: 1, 2 - Charatiopsis, 3 – Centritractus, 4 – ophiocytium
koloniálne, mnohobunkové a nebunkové. Bunková membrána je hustá, pektín a celulóza, pozostávajúca z tesne sa prekrývajúcich častí alebo z dvoch ventilov. V škrupine je uložený oxid kremičitý alebo vápno. Väčšinou fixné formy. Medzi jednobunkovými druhmi existujú mobilné formy, ktoré nemajú hustú škrupinu a sú vybavené bičíkmi, lobopódiami a rizopódiami.

Ryža. 45. Vzhľad xantokokových žltozelených rias: 1–3 - Botrydiopsis, 4 – tetrahedriella, 5 – Pseudostaurastrum, 6 – Goniochloris, 7, 8 – Bumilleriopsis
Najviac žltozelené – nehybné organizmy. U mobilných jedincov sa pohyb môže vykonávať pomocou bičíkov alebo rizopódií. Bunky rôznych tvarov: guľovité, vretenovité, elipsoidné, valcovité, štvorstenné, kosáčikovité, hruškovité, vajcovité. Veľkosti Thallus od 0,5 – 1,5 µm ( Chloridella) do priemeru niekoľkých milimetrov ( Botrydiopsis) (obr. 45, 1 – 3) a dĺžky až do desiatok centimetrov ( Vosheria) (obr. 46, 3).

Ryža. 46. Vzhľad žltozelených rias: 1 - Tribonema, 2 – heteropédia, 3 – Vosheria, časť vlákna s oogóniom a anteridiom
Väčšina žltozelených druhov sú fototrofy, ale nachádza sa aj holozoické kŕmenie požitím baktérií a malých rias. Žltozelené riasy sú široko rozšírené v sladkých vodách. Časté sú aj v pôde, menej časté v morských a brakických vodách. Trieda zahŕňa aerobiontné, planktonické, bentické a perifytonické formy. Epifyty, epizoity, ako aj intracelulárne symbionty v bunkách prvokov.
Bez ohľadu na vonkajšiu štruktúru je vnútorná štruktúra bunky žltozelených rias rovnaká. V protoplaste sa zvyčajne nachádza niekoľko žltozelených chloroplastov, ktoré majú diskovitý, korytovitý, lamelárny, menej často stuhovitý, hviezdicovitý alebo miskovitý tvar s celistvými alebo laločnatými okrajmi. Sfarbenie je spôsobené absenciou fukoxantínu, ktorý je zodpovedný za zlatú a hnedú farbu iných ochrofytov. Z ostatných pigmentov majú β -karotén, vosheriaxantín, diatoxantín, diadinoxantín, heteroxantín. V mobilných formách sa červené oko alebo stigma zvyčajne nachádza na prednom konci chloroplastu. Len málo druhov má pyrenoidy poloponoreného typu. Jadro v bunke je zvyčajne jedno, má malú veľkosť, ale existujú druhy s viacjadrovými bunkami. Niektoré druhy majú jednu alebo dve kontraktilné (pulzujúce) vakuoly v prednej časti bunky.
Zástupcovia monád a pohyblivé štádiá (zoospóry a gaméty) majú dva nerovnaké bičíky. Výnimkou sú synzoospóry. Vosheri, v ktorej sa pozdĺž povrchu nachádzajú početné páry hladkých bičíkov mierne odlišných dĺžky. Krátky bičík končí akronémou. Bičíky sú pripojené k bunke subapikálne. Pri spermiách Vosheri bočné pripevnenie.
U druhov s améboidnou, monadálnou a palmeloidnou organizáciou bunková stena chýba, sú pokryté iba cytoplazmatickou membránou a môžu ľahko meniť tvar. Niekedy sa vo vnútri domov nachádzajú „holé“ bunky, ktorých steny môžu byť natreté hnedou soľou mangánu a železa. Prevažná väčšina foriem má bunkovú stenu pozostávajúcu z dvoch častí. V zložení bunkovej steny prevláda celulóza a obsahuje aj polysacharidy, pozostávajúce najmä z glukózy a urónových kyselín. V mladých bunkách je membrána tenká, s vekom sa zahusťuje. Môžu sa v ňom ukladať soli železa, ktorých zlúčeniny ho farbia do rôznych odtieňov hnedých a červených tónov. Častejšie je oxid kremičitý prítomný v bunkovej stene, čo jej dodáva tvrdosť a lesk. Môže byť tiež pokrytý vápnom a môže byť vytvarovaný rôznymi spôsobmi (tŕne, bunky, bradavice, zubáče, atď.) Výrastok škrupiny sa môže vytvoriť v pripojených formách – chodidlo s pripevňovacou podrážkou.
Vo filamentóznych formách žltozelených rias s lastúrnikmi sa pri rozpade filamentov bunkové membrány rozpadajú na úlomky v tvare písmena H. Tieto fragmenty sú pevne spojené polovice schránok dvoch susedných buniek (obr. 47). Počas rastu filamentov je fragment bunkovej steny dvoch susedných dcérskych buniek v tvare písmena H vložený medzi dve polovice membrány materskej bunky. Výsledkom je, že každá z dcérskych buniek je z polovice pokrytá starou membránou materskej bunky a z polovice – novovytvorená škrupina.

Ryža. 47. Schéma vzniku priečnej priehradky medzi dvoma dcérskymi bunkami vo vláknitých žltozelených riasach (podľa: A.A. Masyuk, 1993): A- fragment vlákna; B- položenie opaskového prstenca a vytvorenie priečnej priečky medzi dvoma bunkami; IN– vrstvenie membrán lastúrnikov; G– rozpad škrupiny na časti v tvare písmena H
V mobilných zástupcoch sú prítomné kontraktilné vakuoly. Zvyčajne sú 1-2 na bunku. Golgiho aparát zvláštnej štruktúry. Diktyozómy sú malé, obsahujú 3-7 cisterien. Jadro je jedno, menej často je veľa; u cenotických druhov sú bunky vždy viacjadrové.
reprodukcie. Väčšina druhov žltozelených rias sa vyznačuje vegetatívnym a nepohlavným rozmnožovaním.
Vegetatívne rozmnožovanie uskutočnené rôzne cesty: delenie buniek na polovicu, rozpad kolónií a mnohobunkových stielok na časti. O Vosheri vytvárajú sa špeciálne plodové puky.
O asexuálna reprodukcia Môžu sa tvoriť rôzne spóry: améboidy, zoospóry, synzoospóry, autospóry, hemizoospóry, hemiautospóry, aplanospóry. Zoospóry sú „nahé“ a zvyčajne majú hruškovitý tvar.
sexuálny proces- izogamia, heterogamia a oogamia - opísané u niekoľkých zástupcov. O Tribonemes gaméty majú podobnú veľkosť, ale líšia sa správaním - ide o izogamiu. O Vosheri pozoruje sa oogamia: na vláknach sa vytvárajú nádobky ženských gamét – oogónia a muž – anteridia.
IN nepriaznivé podmienky pozoruje sa tvorba cysty. Cysty (statospóry) sú endogénne, jednojadrové, zriedkavo viacjadrové. Ich stena často obsahuje oxid kremičitý a pozostáva z dvoch nerovnakých alebo zriedkavejšie rovnakých častí.
Systematika.
Na konci XIX – začiatkom 20. storočia do oddelenia zelených rias boli priradené rôzne rody žltozelených rias, čo súviselo predovšetkým s farebnou a morfologickou podobnosťou stél. Žlté zelené sa v súčasnosti považujú za triedu v rámci divízie ochrofytov.
Známych je asi 450 moderné druhy trieda Žltozelené riasy, ktoré sú zoskupené do štyroch radov: Botridia, Mishococcus, Tribonem a Wocheriaceae. Klasifikácia objednávok je založená na type diferenciácie talu a znakoch životného cyklu.
Rad Botridiae - Botrydiales. Rad zahŕňa druhy so sifonálnym typom diferenciácie talu, v ktorých nie je žiadny oogamný pohlavný proces.
Rod Botridiumžije na pôde a má vzhľad zelených bublín s veľkosťou niekoľkých milimetrov, pripevnených bezfarebnými rizoidmi. Talus je sifonálny, obsahuje početné jadrá a plastidy. Škrupina je viacvrstvová, môže sa na ňu ukladať vápno. Rozmnožovanie je nepohlavné pomocou biflagelátových zoospór, pričom celý obsah močového mechúra sa rozpadá na jednojadrové úlomky. Pri nedostatku vlahy sa rozmnožuje pomocou aplanospór alebo tvorí hrubostenné cysty. V niektorých prípadoch celý obsah močového mechúra ide do tvorby jednej veľkej cysty. V iných prípadoch sa cysty tvoria v rizoidoch, kde najskôr prechádza obsah močového mechúra. Cysty klíčia buď priamo do nového talu, alebo tvoria zoospóry. Sexuálny proces je izo- a heterogamia. Zygota klíči okamžite, bez obdobia pokoja. Bežné a rozšírené druhy v suchozemských biotopoch, vyskytujúce sa pozdĺž brehov potokov, rybníkov alebo na pôdach bez vegetácie.
Objednajte si Mischococcal–Mishococcales. Jednobunkoví alebo koloniálni zástupcovia s kokoidným typom diferenciácie talu.
Rod Charatiopsis zahŕňa jednobunkové pripojené formy. Pri rozmnožovaní vytvára zoospóry, aplanospóry a hrubostenné cysty (obr. 44, 1-2).
Rod ophiocytium(obr. 44, 4) má podlhovasté valcové články, ktoré môžu byť rovné, ohnuté alebo špirálovito stočené a na konci môžu niesť bodec. Bunková stena sa skladá z dvoch nerovnakých častí, z ktorých väčšina z nich podieľa sa na raste buniek, menšia časť je trvalá a má tvar čiapky. Jednobunkové a koloniálne druhy, voľne žijúce alebo pripevnené k substrátu malou stopkou. Rozmnožujú sa zoospórami a aplanospórami, nachádzajú sa cysty. Žijú v sladkých vodách.
Rod Mischococcus vytvára stromovité prirastené kolónie. Vetvenie je dichotomické a tetrachotomické. Bunky sú umiestnené v 2 alebo 4 na vrcholoch hlienových vetiev kolónie. Bunky sú guľovité až oválne, s tenkou alebo hrubou bunkovou stenou. Niekedy je bunková stena lesklá a hnedá kvôli jej impregnácii soľami železa. Mladý jednobunkové organizmy so slizkou, diskovitou základňou, ktorá slúži ako opora na pripevnenie. Po uvoľnení spór sa protoplast materskej bunky zmení na rôsol a natiahne sa, dĺžka sa zväčší 6-krát ako šírka, a tak sa objaví valcovitá noha. Základom stonky sa vždy stáva prázdna bunková stena materskej bunky. Nepohlavné rozmnožovanie zoospórami a autospórami. Autospóry sú pripevnené k hornému okraju hlienovej stopky. Následné bunkové delenia opakujú proces a vytvárajú kolóniu podobnú stromu. sexuálny proces – izogamia. Žijú v malých sladkovodných útvaroch ako epifyty vláknitých rias. Známy v strednej Európe a Ázii.
Rad Tribonemaceae - Tribonematales. Zástupcovia majú vláknitý, multifilamentózny, falošný tkanivový a tkanivový typ diferenciácie talu. Bunkové steny sú buď s prekrývajúcimi sa časťami v tvare H alebo sú plné.
Rod Tribonema- nerozvetvené nite (obr. 46, 1). Bunky sú valcovité alebo súdkovité. Bunková stena sa skladá z dvoch polovíc, ktoré sa nachádzajú s okrajmi na sebe v strede bunky. Mušle sú často vrstvené. Úlomky nití vždy končia prázdnymi polovicami úlomkov mušlí v tvare písmena H, ktoré vyzerajú ako vidlica. V bunkách je niekoľko žltozelených plastidov, nie sú tam žiadne pyrenoidy. Rozmnožovanie je vegetatívne (fragmentácia filamentov), asexuálne (zoospórami a aplanospórami) a sexuálne (izogamia), pričom aplanospóry sú produkované častejšie ako zoospóry. Môže vytvárať akinéty. Žijú v sladkých vodách, kde sa obzvlášť hojne rozvíjajú v chladnom období.
Objednajte si Vosherievye - Vaucheriales. Všetci zástupcovia majú sifonálny talus, oogamný sexuálny proces a synzoospóry.
Rod Vosheria(obr. 46, 3) má talus nebunkovej štruktúry; jeho stélka dosahuje dĺžku niekoľko centimetrov, prichytáva sa k substrátu pomocou bezfarebného rizoidu. V závitoch nie sú žiadne priečky, väčšina talu je obsadená vakuolou a početné jadrá a plastidy sa nachádzajú pozdĺž periférie v cytoplazme. Nite s vrcholovým rastom a zriedkavým bočným vetvením. Priečky sa vytvárajú pri poškodení talu a na oddelenie reprodukčných orgánov. Nepohlavné rozmnožovanie sa uskutočňuje pomocou aplanospór, synzoospór, akinét. Synzoospóry sa tvoria po jednej v zoosporangiu, ktoré je oddelené od vegetatívnych buniek priehradkou na konci vlákna. Zoospóry sú viacjadrové a mnohobičíkové. Sexuálny proces je oogamia. Zygota je pokrytá hrubou škrupinou a po období pokoja vyrastie do nového talu.
Druhy Vosheriširoko rozšírený v sladkých, brakických a morských vodách, ako aj v suchozemských biotopoch. Vyskytujú sa na všetkých kontinentoch vrátane Antarktídy. Tvoria trávovozelené alebo tmavozelené prepletené hmoty - takzvané rohože, hladké, plazivé alebo vankúšovité. Vodné, polovodné, suchozemské formy. Žijú v rôznych biotopoch: moria, ústia riek, ústia riek, slané močiare, mangrovy, potoky, kanály, jazerá, rybníky, orná pôda a močiare.
Významheterokontné riasy
Z oddelenia rias Heterocont majú pre prírodné ekosystémy a pre človeka najväčší význam hnedé riasy.
Hnedé riasy - hlavné zdroj organickej hmoty v pobrežnej zóne morí. Ich biomasa v moriach mierneho a subpolárneho pásma môže dosahovať niekoľko desiatok kilogramov na meter štvorcový. Húštiny hnedých rias poskytujú podmienky na kŕmenie a rozmnožovanie mnohých pobrežných živočíchov a iných rias. Charles Darwin pozoroval hnedé riasy pri pobreží Južnej Ameriky Macrocystis, napísal: „Tieto rozsiahle podvodné lesy južnej pologule môžem porovnať iba s pozemskými lesmi tropických oblastí. A predsa, ak by sa v niektorej krajine zničil les, nemyslím si, že by zahynul ani približne rovnaký počet živočíšnych druhov ako pri ničení tejto riasy.
Poslúžia húštiny hnedých rias miesto na jedlo,prístrešie a chov veľa zvierat. Obrazne povedané, hnedé riasy poskytujú iným vodným organizmom „stôl, úkryt a škôlku“.
Hnedé riasy sú široko používané ľuďmi. Sú bohatí jód a ďalšie mikroživiny. Národy juhovýchodnej Ázie ich tradične používajú pre jedlo, najmä zástupcovia rádu kelu, z ktorých sa pripravuje široká škála jedál. Kŕmiť múku, pripravený z hnedých rias, zvyšuje produktivitu hospodárskych zvierat; zároveň sa vo vajciach a mlieku zvyšuje obsah jódu.
Z hnedých rias prijímaťalgináty- soli kyseliny algínovej. Algináty sú široko používané v rôznych priemyselných odvetviach. Ide o netoxické zlúčeniny s koloidnými vlastnosťami, preto majú široké využitie v potravinárskom a farmaceutickom priemysle. Kyselina algínová a jej soli sú schopné 200-300-násobne absorbovať vodu a vytvárať gély, ktoré sa vyznačujú vysokou odolnosťou voči kyselinám. V potravinárstve sa používajú najmä ako emulgátory, stabilizátory, želírujúce a vodu zadržiavajúce zložky. Napríklad suchý práškový alginát sodný sa používa pri výrobe práškových a briketovaných rozpustných produktov (káva, čaj, sušené mlieko, želé atď.) na ich rýchle rozpúšťanie. Vodné roztoky algináty sa používajú na mrazenie mäsa a rybích výrobkov. Až 30 % z celkového objemu alginátov vyrobených vo svete smeruje do potravinárskeho priemyslu.
V textilnom a celulózo-papierenskom priemysle sa algináty používajú na zahusťovanie farieb a zvýšenie pevnosti ich väzby s podkladom. Impregnácia látok niektorými alginátmi im dodáva ochranné vlastnosti: vodeodolnosť, odolnosť voči kyselinám a zvyšuje mechanickú pevnosť. Na získanie umelého hodvábu sa používa množstvo solí algínových kyselín. Počas druhej svetovej vojny v USA a Anglicku sa z kyseliny algínovej a jej solí vyrábalo veľké množstvo maskovacích látok a sietí pre obytné a priemyselné budovy.
Algináty sa používajú v metalurgii: v zlievárňach zlepšujú kvalitu formovacej zeminy. Pri výrobe elektród na elektrické zváranie sa používajú soli kyseliny algínovej, ktoré umožňujú získať kvalitnejšie zvary. Algináty sa používajú aj pri výrobe plastov, syntetických vlákien, náterov a stavebných materiálov odolných voči poveternostným vplyvom. Používajú sa pri výrobe vysokokvalitných mazív pre stroje. V rádiovej elektronike pôsobia algináty ako spojivo pri výrobe vysokokvalitných feritov.
Najväčšie uplatnenie nachádza vo vode rozpustný alginát sodný, schopný vytvárať viskózne roztoky. Je široko používaný na stabilizáciu rôznych roztokov a suspenzií. Pridanie malého množstva alginátu sodného do potravinárskych výrobkov - konzervy, zmrzlina - zlepšuje ich kvalitu. Vyrába sa z neho aj dekoratívna kozmetika, krémy a masky v parfumérskom priemysle.
Vo farmaceutickom priemysle sa algináty používajú na poťahovanie tabliet, piluliek, ako základ pre rôzne masti a pasty, ako gély na nosiče liečiv a pri výrobe rozpustných chirurgických stehov. V medicíne sa alginát vápenatý používa ako hemostatické činidlo a ako sorbent, ktorý odstraňuje rádionuklidy (napríklad stroncium). Ročná produkcia alginátov vo svete presahuje 20 tisíc ton.
Ďalšou dôležitou látkou získanou z hnedých rias je šesťsýtny alkohol manitol. Manitol sa používa ako náhrada cukru pre diabetikov. Okrem toho sa môže použiť ako náhrada plazmy na konzerváciu krvi. Používa sa na výrobu tabliet vo farmaceutickom priemysle. Manitol sa tiež používa v výroba syntetických živíc, farieb, papiera, výbušnín, apretúry kože.
fukoidany, získané z hnedých rias sú účinné antikoagulanciá, dokonca aktívnejšie ako heparín. Ich použitie na výrobu protinádorových liečiv a antivírusových zlúčenín sa považuje za sľubné. Koniec koncov, aj pri najnižších koncentráciách môžu fukoidany inhibovať prichytenie vírusov na povrch buniek. Fukoidany majú tiež schopnosť tvoriť mimoriadne silný a viskózny hlien, ktorý sa využíva pri príprave stabilných emulzií a suspenzií.
Energetická kríza, ktorá v posledných rokoch zachvátila mnohé krajiny sveta, viedla k potrebe hľadania nových netradičných zdrojov energie. Takže v USA sa na tento účel skúma možnosť šľachtenia obrovských rias. Macrocystis s následným spracovaním na metán. Vypočítalo sa, že z plochy 400 km 2, ktorú zaberá táto riasa, možno získať 620 miliónov m 3 metánu.
Heterokontné riasy z tried Golden, Yellow-green, Sinura, Rafidophyte a Eustigma riasy, zastúpené najmä mikroskopickými organizmami, sú rozšírené v sladkovodných útvaroch všetkých klimatických pásiem zemegule, ale sú bežnejšie v miernych zemepisných šírkach. Medzi zlatými riasami sú druhy, ktoré žijú v moriach a slaných jazerách a veľmi málo v znečistených vodách. Zlaté riasy dosahujú maximálny rozvoj v chladnom období: dominujú v planktóne skoro na jar, neskorá jeseň a zima. V tejto dobe zohrávajú významnú úlohu ako producenti prvovýroby a slúžia ako potrava pre organizmy zooplanktónu.
Napríklad nejaké zlaté riasy Uroglena A Dinobrion, vyvíjajú sa vo veľkom množstve, sú schopné spôsobiť vodné kvitnutie. Uvoľňujú aldehydy a ketóny, ktoré môžu dodávať vode nepríjemný zápach a chuť, a Uroglena– mastné kyseliny toxické pre ryby.
Rafidové riasy sú široko zastúpené v planktóne sladkovodných útvarov s prevažne kyslým pH, najmä v rašelinísk, menej často vo veľkých jazerách. V sladkovodných útvaroch sa môžu vytvárať miestne „kvety“. Goniostomum. Rafidové riasy sa nachádzajú aj v odsolených morských zálivoch a kalužiach na pobreží, ako aj na otvorenom mori. S masovým rozvojom v pobrežných morských vodách spôsobujú toxické "kvety" vody. Takže pri pobreží Kanady počas „kvitnutia“ môže koncentrácia buniek rapídnej riasy Heterosigma dosiahnuť 30 miliónov na 1 liter. Ohniská raphidových rias často vedú k rozvoju „červených prílivov“, ktoré sú spojené s úhynom rýb. Príčinou takýchto "červených prílivov" môžu byť druhy pôrodu Hattonella, Olistodiscus, Heterosigma a Fibrocapsa.
Sinura riasy s hromadným vývojom v sladkovodných útvaroch môžu vode spôsobiť nepríjemný zápach ( Sinura). Riasy Feotamnia sa nachádzajú v stojatých a pomaly tečúcich sladkých vodách, kde sa epifyticky usadzujú na vláknitých riasach.
Riasy Eustigma sa nachádzajú iba v sladkej vode alebo v pôde.
Žltozelené riasy sú rozšírené na všetkých kontinentoch, žijú najmä v sladkej vode a pôde, ako aj v suchozemských, brakických vodách a morských biotopoch. Žltozelené riasy obývajú čisté a znečistené vody s rôznymi hodnotami pH: môžu žiť v kyslých aj zásaditých vodách. Vyskytujú sa hlavne v čistých sladkovodných nádržiach, menej často v moriach a brakických vodách, preferujú mierne teploty, častejšie sa rozvíjajú na jar a na jeseň, hoci existujú druhy, ktoré sa vyskytujú počas všetkých ročných období vrátane zimy. Najčastejšie ich možno nájsť v zhlukoch nití a medzi húštinami vyšších vodné rastliny v pobrežnej zóne riek, rybníkov, jazier a nádrží.
Prevažná väčšina žltozelených sú voľne žijúce formy, ale v bunkách prvokov sú aj vnútrobunkové symbionty – zooxantely. Zaujímavú vnútrobunkovú symbiózu tvoria chloroplasty morských druhov Vosheri s lastúrnikom Elision. Počas deviatich mesiacov je tento mäkkýš schopný fotoautotrofnej fixácie oxidu uhličitého v kultúre. Ide o najdlhšiu symbiózu tohto typu, kedy je symbiotický plastid v priamom kontakte s cytoplazmou zvieraťa. V prírode sa larvy mäkkýšov živia vláknami. Vosheri. V dôsledku fagocytózy sa chloroplasty rias dostávajú do cytoplazmy epitelových buniek mäkkýšov. Počas tohto procesu sa chloroplastová membrána stáva trojvrstvovou, jedna vonkajšia membrána CES (chloroplastové endoplazmatické retikulum) sa stráca. Tento jav je dobrým dôkazom toho, že v priebehu evolúcie by mohli vzniknúť chloroplasty s tromi membránami ako výsledok sekundárnej symbiogenézy v dôsledku straty membrán.
Žltozelené, zlaté a iné heterokontné riasy sú producentmi kyslíka a organických látok; sú súčasťou potravinového reťazca. Heterokontofyty sa podieľajú na samočistení znečistených vôd a pôd, tvorbe sapropelu, na procese akumulácie organickej hmoty v pôde, ovplyvňujúcej jej úrodnosť. Používajú sa ako reprezentatívne organizmy pri určovaní stavu znečistenia vôd; žltozelené riasy sú súčasťou komplexu mikroorganizmov používaných na čistenie odpadových vôd.
Kontrolné otázky
Vymenujte charakteristické znaky štruktúry hnedých rias.
Štrukturálne znaky hnedých rias thalli.
Ako sa rozmnožujú hnedé riasy? Čo sú monospóry, tetraspóry a zoospóry, izogamia, heterogamia a oogamia?
Aké sú životné cykly hnedých rias? Reprodukcia fucus a chaluhových rias.
Vymenujte charakteristické znaky a typických predstaviteľov radov hnedých rias.
V ktorých biotopoch sa nachádzajú hnedé riasy? Aký je ich význam v prírode.
Hospodársky význam hnedých rias.
Vymenujte charakteristické znaky stavby a typických predstaviteľov zlatých rias.
Aké pigmenty a druhy výživy sú známe u zlatých rias?
Rozmnožovanie a ekológia zlatých rias.
Vymenujte charakteristické znaky a typických predstaviteľov žltozelených rias.
Aké pigmenty a druhy výživy sú známe u žltozelených rias?
Ako sa rozmnožujú žltozelené? Typy sexuálneho rozmnožovania: izogamia, heterogamia a oogamia?
Vymenujte charakteristické znaky stavby a typických predstaviteľov sinurických rias.
Rozmnožovanie a ekológia sinurických rias.
Vymenujte charakteristické znaky stavby a typických predstaviteľov rias feotamnia.
Rozmnožovanie a ekológia rias feotamnia.
Vymenujte všeobecné znaky stavby a typických predstaviteľov rafidofytných rias.
Rozmnožovanie a ekológia rafidofytných rias.
Aké sú spoločné znaky štruktúry a typických predstaviteľov rias Eustigma.
Rozmnožovanie a ekológia rias Eustigma.
Hodnota heterokontných rias v prírodných ekosystémoch.
MORSKÉ RIASY
všeobecné charakteristiky
Riasy sú veľkou skupinou nižších rastlínny - od mikroskopicky malých, jednobunkových organizmov až pomnohobunkových obrov. Pojem "riasy" nie je systémmatematická jednotka. Pod názvom "riasy" spájaniekoľko systematických delení nižších rastlín rôzneho pôvodu, charakterizovaných podobným spôsobomživot a autotrofný typ výživy. Ako už názov napovedá,riasy sa zvyčajne vyznačujú vodným životným štýlom.
Telo rias, podobne ako ostatné nižšie rastliny, jestélka - stélka, nerozrezaná na koreň, stonkaa list a u väčšiny druhov má jednoduchú anatomickú stavbu. Riasy obsahujú vo svojich bunkách chlorofylfotosyntetizovať. Štruktúrabunky, telá, spôsoby rozmnožovania sú u jednotlivých zástupcov rôznezástupcovia tejto skupiny závodných divízií.
Bunky väčšiny rias majú celulózovú membránu; existujú bunky, ktorých obsah je obklopený len tenkou hraničnou membránou. Bunkové steny rias sú rôznorodé v štruktúre a chemické zloženie. Základ škrupinyje proteín- sacharidový komplex. Škrupina sa vyznačuje heterogenitou, vrstvením. Vrstvy sa navzájom líšiahrúbka, hustota a chemické zloženie. Často sú škrupiny impregnované Organické zlúčeniny(lignín a kutín).
V škrupine sú špeciálne otvory - póry. Okrem pórov sú bunkové membrány mnohých rias vybavené rôznymi druhmi výrastkov.stami - štetiny, ostne a šupiny.
V protoplaste bunky sa rozlišuje jadro a cytoplazma. Robiť bolesťVäčšina rias v bunke má iba jedno jadro, ale niekedySú ich 2, 3 alebo viac. Bunkám modrozelených rias chýba dobre vytvorené jadro. Tvar, veľkosť a umiestnenie jadra v bunkesa medzi rôznymi riasami značne líšia. V jadre riastvoria sa rovnaké štruktúry ako v jadrách iných rastlín: škrupina,jadrová šťava, jadierko, inklúzie chromatínu.
Cytoplazma pozostáva z hlavnej látky (stroma) a teliesok (organel), ktoré sú v nej ponorené. Charakteristická vlastnosť lepidlaprúdom rias je slabý vývoj endoplazmatického retikula.
V špecifických organelách - chloroplastoch (nazývaných chromomatofóry) sú telá bohaté na bielkovinové látky,nazývané pyrenoidy. Pyrenoid je obklopený podšívkou vo forme prstenca alebo jednotlivých dosiek, zvyčajne škrobovýchprírody. Chloroplasty obsahujú zelený pigment chlorofylexistujúci v niekoľkých formách.
Okrem chlorofylu obsahujú riasy ďalšie pigmenty, ktoréktoré často svojou prítomnosťou maskujú zelenú farbu vody.vyrásť. Najcharakteristickejší a najčastejšie sa vyskytujúci prasiatkariasy, okrem chlorofylu (zelené), karoténu (žlto-oranžový) a xantofyl (žltý), sú fykocyanín, fyko-erytrín a fukoxantín. Phycocyan pigment, rozpustný vo vode,farby rias modrá (modrozelené riasy);fykoerytrín, tiež rozpustný vo vode, dodáva červenú farbu(červené riasy); fukoxantín spôsobuje hnedú farbu(hnedé riasy).
Riasy, ktoré majú pigmenty, dávajú substrát, na ktoromusadzujú sa, rôzne sfarbenie. Napríklad v Antarktíde objavili sovietski vedci 3 jazerá s rôznymi farbamivoda: modrá, zelená a červená. Farba vody týchto jazier bola spôsobenáprítomnosť v nich veľmi malých rias zodpovedajúcej farby.Ľadové brehy Grónska na mnohých miestach s nástupom jarijeho otepľovanie sa stáva červenou (krvavou) farbou, čo sa vysvetľuje hromadným rozmnožovaním rias s červenou farbouhanblivý.
Názov Červené more je spôsobený jeho rastom vo veľkommnožstvo rias v tomto mori, ktoré majú červenkastý odtieň.Rôzne farby rias majú adaptačnú hodnotunie. Denné svetlo až po riasy ponorené hlboko vo vodevždy prichádza v upravenom zložení. Voda je relatívne dobrá.prenáša modré a zelené a silne absorbuje červené a žltélúče. Vo veľkých hĺbkach zadržiavanie zelených lúčov o jedenchlorofyl je zlý, na záchranu prichádza červenápigment, ktorý ľahko absorbuje zelené lúče. Preto červenáalebo karmínové, riasy sú bežné vo veľkých hĺbkachmoriach.
Riasy sú veľmi rozmanité v tvare a veľkosti. Zoznámte sa-Existujú mikroskopicky malé a dosahujúce obrovské veľkosti (až 50 m a viac) jednobunkové a mnohobunkové formy. Prechodné štádium medzi nimi zaberajú koloniálne riasy.Kolónie pozostávajú z niekoľkých voľne spojených homogénnychbunky. S nárastom veľkosti rias, niektorérajská diferenciácia ich tiel. Napríklad v hnedých riasach,rastúce veľké, talus silne rozrezaný. Takéto riasy sú pripevnené k substrátu tenkými bezfarebnými vláknami,ktoré sa nazývajú rizoidy.
Existujú jednobunkové riasy, do ktorých telo zasahujeveľké veľkosti 0,5 m a viac. Majú veľké množstvojadrá a chromatofóry. Vonkajšie má telo takejto riasy rozrezanéale vnútri nie sú žiadne priečky, t.j. je jednobunkový. Príkladom takejto obrovskej jednobunkovej riasy ježivá morská caulerpa.
Reprodukcia rias môže byť vegetatívna, asexuálna a sexuálna. Vegetatívne rozmnožovanie sa vykonáva po častiachtelá rias. K nepohlavnému rozmnožovaniu dochádza v dôsledku tvorby zoospór, menej často spór. Sexuálny proces v rôznych typochriasy zastupujú: izogamia, heterogamia, oogamia aautogamia.
V niektorých viac organizovaných riasach (červené,hnedá) dochádza k striedaniu sexuálnych a nepohlavných generácií.
Riasy sa vyznačujú veľkou rozmanitosťou. generálpočet ich druhov je nad 20 tisíc.Klasifikácia rias je veľmizložité a ešte nie je dokončené. Druhy sa spájajú do väčších systematických celkov – rodov, čeľadístva, zákazky, triedy, oddelenia (typy).
Celá škála rias sa zvyčajne kombinuje do 6 ... 10 veľkých oddelení (typov), ktorých zástupcovia sa líšia štruktúrouny, spôsob rozmnožovania, pôvod, ale najčastejšie predZástupcovia týchto oddelení sa navzájom líšia farbou.
V tejto učebnici sa uvažuje o 6 oddeleniach rias: modrá-zelená, mnohobičíková, rozsievky, zelená, hnedá a červenááno.
Riasy majú veľmi starý pôvod, v ktoromveľa je stále nejasné. Neexistuje jediný uhol pohľadu na vzťah medzi divíziami rias. Niektorí z ich zástupcovočividne pochádzajú z jednoduchších organizmov, ako sú bičíkovce, a samotné riasy sú predkami už dávnejšieskrútené rastliny (huby, machy).
Predstavitelia moderného rozmanitého sveta riasmajú odlišnú antiku vo svojom pôvode, objavili sa v rôznych geologických epochách. Najstaršie súmodro-zelené riasy.
Evolúcia rias prešla od mobilných k nehybným formám.Zvažujú sa primitívnejšie a starodávne skupiny riastí, ktorí trávia svoj život v mobilnom štáte; viac organiNazývané riasy sa vyznačujú ustáleným spôsobom života. Cestaevolúcia rias prišla z prvokov - jednobunkových mikrood skopicky malých foriem až po mnohobunkové komplexné formy.
Ako už bolo spomenuté, drvivá väčšina riasžiť vo vodnom prostredí. Niektoré z nich rastú v slaných vodách oceánov.a moria, iné - v riekach, rybníkoch a sladkovodných jazerách. Bolesťvplyv na distribúciu rias v hĺbkach vodných útvarovposkytuje svetlo, preto v povrchových vrstvách vody, množstvorias je vždy viac. Morské riasyčervená a hnedáčasto tvoria obrovské podmorské húštiny, ktoré pokrývajú desiatky kilometrov. V závislosti od biotopu, riasysa delia na 2 veľké skupiny: bentické a planktónové.
Bentické alebo spodné riasy žijú prichytením saspodné alebo podvodné predmety a tvoria hlavne húštinyv pobrežnej zóne. Planktónové riasy žijú vo vodezavesený stav, nie sú pripevnené na dne, môžu sa prenášaťpohybovať sa pohybom vody.
Riasy žijú nielen vo vode, ale nachádzajú sa aj na povrchupôde, v pôde, na stromoch, na hromadách, ale vždy vlhkáMiesta. Často, najmä skoro na jar, pôdy "kvitnú",alebo "zozelenať", čo sa vysvetľuje vývojom kolosálnych colimikroskopicky malé riasy. „kvitnutie“ pôdy zriasy sa vyskytujú v rôznych oblastiach Sovietsky zväz - v severnej, stepi a dokonca aj púšti. Rastúce do vo veľkom počte, vytvárajú podmienky pre rozvoj baktérií a inémikroorganizmy.
Riasy majú veľký význam v prírode a široko používanýsú volané osobou.
Ako autotrofné rastliny spracúvajú obrovské colimnožstvo minerálov a oxid uhličitý do organickej hmoty. Riasy vytvárajú pre zviera obrovské zásoby potravysvet morí, oceánov a sladkovodných nádrží. Takže 1 ha húštiny rias môže priniesť výnos rovný 100 tonám surovej alebo 10 tonám suchejomši.
Riasy produkujú obrovské množstvo kyslíka.
Rozvoj rybolovu je neoddeliteľne spojený s riasami. Tým, že absorbuje veľa oxidu uhličitého a uvoľňuje kyslík, riasyvyčistiť nádrže.
V prímorských krajinách (Anglicko, Francúzsko, Nórsko, Írskoatď.), najmä v Japonsku sú riasy široko konzumované(morské riasy atď.) a na krmivo pre hospodárske zvieratá (v surovej, suchej a silážovanej formekúpeľňa). Niektoré riasy sa používajú ako hnojivopoliach.
Mnohé riasy akumulujú veľké množstvo jódu a brómu. Jód je obsiahnutý v popole rias asi 0,2% sušiny.Riasy sú často hlavným zdrojom jódu. Trávením červených a hnedých rias získate cennú látku - agar-agar,ktorý sa používa v mikrobiológii ako živná pôdana kultiváciu mikroorganizmov, ako aj v cukrárnipriemysel pri výrobe marmelád a pod.
V starovekých geologických obdobiach boli rozsievky spolus oxidom kremičitým vytvorila sedimentárna hornina – tripol, ktorápoužíva sa pri výrobe dynamitu, tehál, na lešteniepoložky atď.
V sladkovodných útvaroch sa riasy podieľajú na tvorbe sapropula, alebo organický kal. Sapropel obsahuje veľké množstvov organickej hmote a často sa používa na bahennú terapiu.Sapropel, ktorý obsahuje veľké množstvo vápenatých solíión, fosfor, železo, možno použiť na poľnohospodárske krmivohospodárske zvieratá.
Riasy, najmä morské, môžu byť škodlivé aj vtedyvo veľkých množstvách pokrývajú podvodné časti lode. Ohromadné odumieranie rias spôsobuje škody vodou a asnásledne úhyn rýb.
Rozdelenie modrozelených rias
Modrozelené riasy alebo kyanid súnajprimitívnejšieho a najstaršieho pôvoduorganizmov. Na základe paleobotanických údajovJe pochmúrne, že moderné modrozelené riasy sa príliš nelíšiaod svojich fosílnych predkov. Vo väčšine prípadov sú zastúpené jednobunkovými formami, hoci existujú aj mnohobunkovénye formy zhromaždené v kolónii.
Ako ukazuje samotný názov tohto oddelenia rias, pre nichcharakteristická modrozelená farba rôznych odtieňov v závislosti odmostíky na pomere pigmentov - chlorofyl, karotén, fyko-cyanín a fykoerytrín.
Bunková membrána modrozelených rias pozostáva z pektínuvyh látky a je z vonkajšej strany pokrytá slizovitou substanciou. V ich klietkekah nemá morfologicky odlišné jadro a chromatofóry,cytoplazma je impregnovaná pigmentmi, a preto sa nazýva chromatoplazma.
V procese asimilácie namiesto obvyklého rastlinného škrobu,vzniká malý sacharidový glykogén (živočíšny škrob).
Modrozelené riasy sa množia veľmi rýchlo, s jednoduchýmbisekcia buniek. Proces ich sexuálnej reprodukcieneprítomný.
Podľa štruktúry a povahy bunkového delenia modrozelenej vody-výrastky sú podobné baktériám. Oni, rovnako ako baktérie, nemajú jasne definované jadro, nemajú žiadne bičíkové štádiá vývoja.existujú. V niektorých modrozelených riasach (vláknitých)Redukcia prebieha v oblastiach, do ktorých sa tieto riasy rozpadajú. Takéto oblasti sa nazývajú hormogónia. Pri nepriaznivomza príjemných podmienok sa z obyčajných buniek tvoria spóry, ktorépokrytý hrubým plášťom. Chráni obsahpred nepriaznivými podmienkami, vďaka čomu sa riasy zadržiaživotaschopnosť po dlhú dobu. Na začiatkupriaznivé podmienky, spóry klíčia a dávajú vznik novémubunka.
Modrozelené riasy žijú hlavne v sladkej vode.vody - rybníky, jazerá, rieky, ale vyskytujú sa aj v moriach,na povrchu pôdy, na skalách. Modré zelené riasy môžužiť ako keby nízke teploty na snehu a ľade a vo výšketeploty (do 80 °C) v horúcich prameňoch. Po smrtibunky hmota rias vo forme vločiek špinavo zelenkavej farbypláva na hladinu vody.
Staroveký pôvod, nediferencovaná štruktúra lepidlaprúd, absencia vytvoreného jadra a pohlavného procesu, rozmnožovaniejednoduchým delením buniek schopnosť vytvárať spóry -všetky tieto znaky poukazujú na primitívnosť modrozelenejriasy. svojim spôsobom zjednodušenék určitej štruktúre sú výrazneodlišné od ostatných riasOna a jej najbližší stoja pred baktériami.
Modrozelené riasy majú dobrú prispôsobivosťodolnosť voči rôznym podmienkamjamy životné prostredie, ktorý jeprispeli k ich zachovaniuaž do súčasnosti bez veľkého množstvazmeny.
Delenie modrozeleného vodíkasli spája asi 1400 widov. Takouto vodou môžu byť zástupcovia tohto oddeleniarástli ako chroococcus, oscilatoria, nostoc atď.
Chroococcus- jednobunková riasa Sharovýrazná forma, niekedy tieto riasy vytvárajú akoby kolónie.Často sa takéto riasy zhromažďujú v skupinách po 2 ... 4, oddelenétenká priehradka a obklopená bežným dosť hustým hlienomhrubá vrstva.
Chroococcus je široko rozšírený v močiaroch medzi vodné rasy.tiene, medzi hrbolčekmi a blatom.
Oscilačné- vláknitý modrozelenýriasy, bežné v stojatých vodách, častovytvára na hladine vody alebo na bahnitom dne tmavozelený film. Bunky tejto riasy majú valcovitý tvar.spojené v jednom vlákne.
Nostok - vláknité modrozelené riasy, vláknaalebo jeho reťazce sú spojené do kolónií, často guľového tvaru,veľkosti plodu slivky. Vonku sú tieto kolónie pokryté želatínouomša. Nostok žije pozdĺž brehov rybníkov a jazier, na vlhkej pôde.a na dne nádrží.
Spolu s hubami sa tvoria niektoré modrozelené riasyrôzne druhy lišajníkov.
Oddelenie zmiešaných väzov morské riasy
Multiflagellate alebo žltozelená,riasy sa vyznačujú tým, že ich zoospóry majú2 nerovnaké bičíky, pričom krátky bičík je hladký a dlhý bičík perovitý. Chromatofory žltozelené, diskovité.Jednobunkové, koloniálne, vláknité a nebunkové organizmy.Príkladom je botrídium.
Botridium- najtypickejšípestovateľ mnohobičíkových rias. Talus je zelená vezikula s priemerom 1-2 mm na spodnej strane vezikulysú umiestnené bezfarebné rozvetvené výrastky - rizoidy, ktorériasy sú zapustené v pôde. Je to jednobunkový mnohojadrový útvarmorské riasy. Stred vezikuly je naplnený bunkovou šťavou, cytoplazma je umiestnená parietálne. Chromatofory obsahujú veľa karotinoidy, preto má botrídium žltozelenú farbu.Pyrenoidy chýbajú.
Rozmnožuje sa hlavne zoospórami, ktoré vznikajúv kolosálnom množstve uprostred bubliny. Žije vo vlhkuzem, pozdĺž okrajov kaluží, tvorí na pôde tmavozelené nájazdyfarby.
Oddelenie rozsievok morské riasy
Rozsievky, rozsievky, alebo oxid kremičitý, alebo bacily, riasy sú extrémnečajová odroda, mikroskopicky malá, väčšinouprípady jednobunkových organizmov. Toto oddelenie rias je charakteristickérizovaetsya zvláštna štruktúra buniek. Bunka je pokrytá pevnou látkouškrupina vo forme pektínu, želatínového filmu, vonkuje pokrytá pazúrikovou škrupinou, ktorá pozostáva z dvoch jastojace polovice, takzvané šerpy. Jeden z týchtoskala pokrýva druhú ako veko na krabici. Pozdĺž poloviceškrupina má na oboch stranách štrbinovitý otvor. Skrz tootvorenie cytoplazmy bunky komunikuje s vonkajším prostredím. cieľki majú výnimočnú silu. NetráviaZvieratá a vtáky Xia sa nezničia ani pri žiarenív plameňoch. Existuje viac ako 5000 druhov. Výrazné názoryHlavnými znakmi sú tvar buniek a rôzne zahustenia naškrupiny vo forme jaziev, sietí atď.
Tvar buniek určitých druhov rozsievok jepredĺžený štvorcový, eliptický, okrúhly, hviezdicovitý, v tvarestuhy, špirály a pod. Bunky obsahujú cytoplazmu, jadro a jedenalebo viac chromatofórov. Okrem chlorofylu a fukoxantínu,Chromatofory obsahujú aj iné žltohnedé pigmenty,preto sú chromatofóry žlté. V bunkách rozsievok nie je škrob, rezervné látky sú zastúpenéľanový olej.
Rozmnožujú sa najmä priamym delením, ktorékráčajú svojou vlastnou cestou. Počas delenia každá dcérska bunkaprijíma jadro, jeden chromatofor a iba jeden z chlopní plášťa,druhý záhyb sa vytvorí nanovo. Okrem priameho delenia, diatomorské riasy sa pri splynutí rozmnožujú sexuálnedve bunky, ktoré predtým zhodili škrupinu.
Rozsievky žijú v morských a sladkých vodáchčasto sú hlavnými neoddeliteľnou súčasťou planktón a slúžia ako cenná potrava pre zvieratá. Listy odumretých buniek rias klesajú ku dnu a postupne vytvárajú obrovské nánosy,známy pod názvom „horská múka, diatomit, tripoli.
Veľké ložiská rozsievok vo forme rozsievok ala sú sústredené v polárnych oblastiach oceánov, blízko Aljašky,Aleutské ostrovy, v Okhotskom mori a Beringovom mori. Dostupnésú v Baltskom mori.
Medzi rozsievky patrí sladkovodná riasa pineulusria,fragilariaa vysvedčenieria; tieto riasy tvoria kolónie vo forme stužiekalebo reťaze;navicularastie huňatá coloniei.
Oddelenie zelených rias
Zelené riasy sú jedným z najrozmanitejších oddelení rias, združuje asi 5000 druhov.Zástupcovia tohto oddelenia sa vyznačujú zelenou farbou,ktorý je spôsobený chlorofylom a nie je maskovaný žiadnymalebo iné pigmenty. Zelené riasy sú zastúpené jednobunkovými, mnohobunkovými, koloniálnymi formami. Zeleníriasy majú najčastejšie vláknitú štruktúru, vlákna pozostávajú zjeden riadok buniek. Bunka má celulózovú membránuplazma, jadro a chromatofóry. Reprodukcia prebieha asexuálnevegetatívne a sexuálnezelené riasy žijú vo vode,ale niektorí z nich žijú aj na súši, natémy.
Prítomná bude väčšinailey žiť ďalejstromy, na kamenenyah na vlhkých, tienistých miestach. V rybníkoch a riekach tvoriabahno.
Oddelenie zelených rias je rozdelené do niekoľkých tried, z ktorýchpovažujme za rovnocenné bičíky, alebo vlastne zelené, páriace sa riasy alebo konjugáty, a char, alebo lúčeTrieda equiflagellate Medzi zelenou vodourastú, táto trieda je najrozsiahlejšia. Zástupcovia sú rôznorodívo vzhľade a vnútornej štruktúre. Sú charakteristicképrítomnosť dvoch identických bičíkov Táto trieda kombinuje jedenbunkové a koloniálne, mobilné a imobilné formy.Trieda je rozdelená do 8 rádov. Zoberme si zástupcov 4 podľariadkov.
objednať volvox . Väčšinatypickými predstaviteľmi radu Volvox súchlamydomonas a volvox.
Chlamydomonas je jednobunkovýmobilné morské riasy. Žije vo veľkom počtev sladkých plytkých vodách - kaluže, rybníky, priekopypozdĺž ciest. Pri hojnom rozmnožovaní zafarbuje vodu na zelenofarba. Je to mikroskopicky malá oválna bunkaalebo okrúhleho tvaru. Na jednom (prednom) konci je klietka predĺženávo forme výlevky, na rovnakom konci sú 2 rovnaké bičíky, ktoré prispievajú k pohybu rias. Celá dutina bunky je vyplnenánie je pokrytá cytoplazmou, v ktorej sa nachádza jadro, bližšie k nosu
Volvoks, príprotujúca vrchná časť, je charakteristickým príkladom koloniálnych foriem mikroskopických rias. Kolónia tejto riasy je viditeľná voľným okom, dosahujeveľkosť špendlíkovej hlavičky a má tvar gule. Takáto sférická kolónia Volvox pozostáva z obrovského počtu buniek (až 50 tisíc), umiestnených v jednej vrstve pozdĺž obvodu gule.Každá bunka nesie 2 bičíky. Všetky bičíky sú umiestnené pozdĺžperiférii a uľahčujú pohyb celej kolónie. Pohybbičíky všetkých buniek sú vždy koordinované. Dutina lopty je vyplnenánepokryté tekutým hlienom. Volvox sa rozmnožuje vegetatívne asexuálne. Počas vegetatívneho rozmnožovania v matkekolónia (guľa) Vznikne 8 ... 15 dcérskych kolónií (guliek). Odozrievaním dcérskych kolónií sa steny dospelého klbka trhajúa vychádzajú mladé kolónie, po ktorých materské kolonia zomiera.
Pohlavné rozmnožovanie – oogamia – sa vyskytuje len u väčšinyzávetria väčšie bunky. Zároveň v bunkách kolónie Volvox, ccaholé dvojbičíkové gaméty sú oddelené, spájajú sa iba gaméty z rôznych jedincov riasy.
Existuje mnoho typov Volvox. Všetky sú rozšírenéhlavne v stojatej, dobre ohriatej sladkej vodevodné nádrže so stojatou vodou – rybníky, jazerá, menej často v riekach
Chlorellamikroskopické jednobunkovéguľovité morské riasy. Chlorella sa vyznačuje rýchloureprodukciu a veľmi aktívny proces fotosyntézy. Vďakaprítomnosť veľkého množstva plastidov chlorelly má odlišný účinokaktívne používanie solárna energia, konvenčné kultúrnerastliny spotrebujú len 0,1 % slnečnej energie s biochemickouikálne premeny a chlorella -2,5%, t.j. 25-krát viac.Iné pozitívna kvalita chlorella - veľmi rýchly rastzhenie.
Ulothrix je vláknitá, nerozvetvujúca sa riasa,pozostávajúce z jedného radu buniek. Spodná bezfarebná závitová bunkamá zvláštny pretiahnutý tvarPomocou tejto bunky sú riasy pripevnené k podvodným predmetom.
Vlákno ulotrix sa predlžuje v dôsledku priečneho delenia bunieka môže rásť donekonečna. Bunky sú homogénnekrátke, každá z nich cytoplazma, jadro, chromatoformys pyrenoidmi. Ulothrix sa rozmnožuje hlavne nepohlavne.vytvorením 4-bičíkatých zoospór. Tieto zoospóry súpresunúť čas a potom sa usadiť na nejakom podvodnom objektea prerásť do nového vlákna.Sexuálny proces je izogamia. Navyše mužské a ženské gaméty sú navonok nerozoznateľné, ale fyziologicky sú rozdielne a pochádzajú z rôznych vlákien, teda rozdielnegaméty sú označené + a -.
Ulothrix žije v sladkej vode, kde rastievodné predmety (kamene, hromady), ktoré sa sfarbujú do jasne zelenej farbyfarba.
Cladophora je vláknitá zelená riasa.Talus pozostáva z rozvetvených nití tvorených jedným radommnohojadrových buniek, jednotlivé rastliny vyzerajú ako pod vodoukríky Bunky sú veľké, mnohojadrové, s hustým,vrstvená škrupina. Nachádza sa v sladkých a slaných vodách
Divízia červená riasy, alebo šarlátové naschiExistuje viac ako 600 rodov a asi 4000 druhov. z iných riassa líšia sfarbením, čo je spôsobené prítomnosťouokrem chlorofylu majú ešte dva pigmenty – fykoerytrín (červený) a fykokyán (modrozelený). Z pomeruFarba rias závisí od tvorby týchto pigmentov (líši sa odčervená až takmer čierna). Zástupcovia oddelenia v prevažnej väčšine mnohobunkovýchnye organizmy komplexná štruktúra a len tie najprimitívnejšiemajú jednobunkový alebo koloniálny talus. Častejšie talusmá podobu kríkov, tanierov, u niektorých je veľmi silne členitý.
Červené riasy žijú v moriach vo väčších hĺbkach akozelená a hnedá. Je to kvôli červenému pigmentuprispieva k zachyteniu zelených a modrých lúčov vo veľkých hĺbkachspektra a tým zlepšuje proces fotosyntézy. V červenej farbeusadenina z rias nie obyčajný škrob. Na rozdiel odobyčajný škrob, nefarbí sa jódom na modro, ale na červenohnedá farba.
Červeným riasam chýbajú zoospóry a spermie.Asexuálna reprodukcia v nich prebieha pomocou imobilnýchspor. Sexuálna reprodukcia - oogamia, ale namiesto spermiívznikajú samčie bunky - spermie, do vajíčok sa prenášajú vodou. Oocyty sa tvoria v špeciálnych orgánoch -karpogóny.
_Červené riasy zahŕňajúdelasseries talom vo forme kríka; polysifóniav ktorom má slez tvar rozvetvenej nite.
oddelenie hnedá morské riasy
Pre zástupcov oddelenia hnedých riastrnka hnedá farba v dôsledku prítomnosti v chromatofórochhnedý pigment - fukoxantín. Prítomnosť fukoxantínumaskuje zelenú farbu a dodáva týmto riasam hnedú farburôzne odtiene. Okrem fukoxantínu obsahujú xantofylo a karotén. Oddelenie hnedých rias sa spája900 druhov.
Hnedé riasy sa zvyčajne vyznačujú veľkým mnohobunkovým talom. Najväčší zástupcovia rias sa nachádzajú práve medzi hnedými riasami. Niektoré z nich, ako makrocystis, dosahujú dĺžku 60 m, ale existujú aj malé formyniekoľko milimetrov od seba.
Namiesto škrobu obsahujú bunky hnedých rias glukózupre a cukrové látky - láka a kelp, ktoré dávajúTieto riasy majú pri varení sladkú chuť. Akorezervnú látku často ukladajú oleje.
Talus hnedých rias je trvácny, ale listnatýplatne každoročne odumierajú a na začiatku jari opäť rastú.
Zložitá vonkajšia štruktúra hnedých rias určujepriich a diferenciácia v anatomickej štruktúre (majúrôzne tvary buniek). Niektorí vedci sa domnievajú, že tietoriasy majú dokonca rôzne tkanivá.
Hnedé riasy sa rozmnožujú rôznymi spôsobmi. Nekoniektoré z nich sa rozmnožujú primitívnymi sexuálnymi prostriedkami – isomia, keď sa spoja 2 gaméty rovnakého tvaru. iné,vyvinutejšie riasy (kelp) tam je zložitejšiepohlavný proces - oogamia, pri ktorej je veľké vajíčkosplýva s malou pohyblivou samčou gamétou - spermiamidom.
V hnedých riasach dochádza k nepohlavnému rozmnožovaniuzoospóry, ktoré sa v zoospo produkujú vo veľkom množstvehodnosti. V hnedých riasach je striedanie generácií celkom jasne vyjadrené; asexuálne a sexuálne. Na listových čepeliachtieto riasy tvoria jednobunkové zoosporangiá, zbieranénye do skupín, medzi ktorými sú neplodné nite.Každé zoosporangium produkuje 16...64 alebo viaczoospóry. Zoospóry sú navonok identické, ale fyziologicky odlišné.Niektoré z nich vyklíčia a vytvoria mikroskopicky malé manželky.skye, a ďalšie - mužské gametofyty. na samčie gametofytyvznikajú ďalšie anterídie a v nich jedna spermiazoid, a na samičích gametofytoch sa tvoria oogónie, nesúcejedno vajce. Po fúzii spermií a vajíčkavzniká zygota, z ktorej sa vyvíja nepohlavná generácia -sporofyt.
Hnedé riasy sú morský život, mnohé z nichsú bežnejšie v severných moriach, často sa tvoriaobrovské húštiny v moriach a oceánoch. Severne od Atlantioceáne, v Sargasovom mori, v obrovskom počteExistuje rod hnedých rias - Sargassum. Týchto rias je viacvšetky sú v plávajúcom stave kvôli prítomnostišpeciálne bubliny naplnené vzduchom.
Hnedé riasy sú považované za starodávnu skupinu rastlín, majúvyšší stupeň diferenciácie sa nachádza nielenvonkajšej, ale aj vnútornej časti talu. Navonok sú podobnéshimi rastliny, takže niektorí botanici veria, že tietoriasy mohli viesť k vzniku vyšších rastlín.
Rozdelenie hnedých rias pozostáva zo 4 rádov. Zvážte predtýmputter dvoch rádov: kelp a fucus.
Objednajte si kelp. Totoveľmi veľké riasy, niekedy dosahujúce 60 m alebo viac.Ich slez je silne členitý a navyše dobre vyvinutýrozvetvené rizoidy, ktorými sú riasy pevne prichytené na dne mora. Laminaria žijú v pobrežnom páse morív hĺbke 5 ... 10 m a často tvoria obrovské húštiny pod vodou„lesy“.
Medzi lamináriá patrí rod Laminaria (zahŕňa 30 druhov), rod Lessonia (zahŕňa5 druh) a rod Macrocystis. Tieto riasy sú viacročné rastliny, ktoré sa navzájom líšiaštruktúra talu.
oddelenie slizové formy
Plasmodium vzniká splynutím nahých amébrôznych buniek slizoviek a v niektorých typoch slizniakov dosahujeveľkosť ľudskej dlane.On zvyčajne sfarbené do jasne žltej farbyfarba a má schopnosť veľmi pomaly pripomínať amébupohyb (0,1 mm/min). Pri pohybe má plazmodium tendenciuskryť sa pred svetlom a vydať sa smerom k zdroju vlhkosti. Zoznámte saslizniaky sa zvyčajne vyskytujú v tienistých lesoch; na hnilých rastlinách, medzikôry a dreva, v popraskaných pňoch, pod opadaným lístím.Rozmnožujú sa spórami.
Pozícia slizniakov vo fylogenetickom systéme je nejasná,zrejme pochádzajú z nejakého druhu bičíkov. Odtiaľtooddelenie, zvážte plazmodiofór.
Choroba sa šíri pôdou a rozvíja sa obzvlášť rýchlo.darí sa mu na kyslých pôdach.
Zdá sa, že všetky heterokonty majú bičíkový aparát. Bičíky sú 2 a na jednom z nich sú veľmi charakteristické rúrkovité trojčlenné perovité výrastky alebo chĺpky - mastigonémy. Práve prítomnosť dvoch nerovnakých bičíkov slúžila ako základ pre pomenovanie tejto obrovskej skupiny vodných organizmov. multiflagelatedriasy, alebo heterocontami. Okrem rozdielov v dĺžke sa bičík líši aj morfologicky: hlavný bičík pozostáva z osi a na nej perovito umiestnených brvitých chĺpkov, bočný bičík je hladký a bičíkovitý. Podobné je aj zloženie pigmentov – chlorofylov A A s a početné karotenoidy, ako aj štruktúra chloroplastov. Chloroplasty sú obklopené štyrmi membránami, pričom dve vonkajšie membrány sú spojené s endoplazmatickým retikulom bunky.
V rámci oddelenia Heterocont (Diverzifikovaných) rias je niekoľko skupín rias zjednotených v triede: Hnedá, Zlatá, Žltozelená, Rafidophytes, Sinura, Feotamnia, Eustigma, niektoré boli predtým považované za samostatné oddelenia.
Trieda Hnedé riasy– Phaeophyceae (Fucophyceae)
Názov triedy „hnedé riasy“ odráža farbu ich talu (z gréčtiny. Rhaios- hnedý). Farba je spôsobená prítomnosťou veľkého množstva hnedých a žltých pigmentov - okrem chlorofylov A A s A β -karotén v ich chloroplastoch je prebytok xantofylov, najmä fukoxantínu.
Mnohobunkové, prevažne veľké riasy, rozšírené najmä v studených vodách oboch hemisfér. Medzi hnedými riasami nie sú známe ani jednobunkové ani koloniálne formy. Je známych asi 1,8 tisíc druhov.
Bunková štruktúra. Bunka u hnedých druhov rias je eukaryotická: vytvorila organely s vlastnými membránami: jadro, mitochondrie, plastidy atď. Zelené pigmenty - chlorofyly A A s- maskované ďalšími: oranžovými karoténmi a žltohnedými xantofylmi; z posledne menovaných prevláda fukoxantín. V bunkách hnedých rias sa nachádza jeden až veľa plastidov. Na povrchu vonkajšej bunkovej membrány bunky hnedých rias zvyčajne vytvárajú tuhú bunkovú stenu.
Jadro je najčastejšie jedno, ale niekedy je jadier niekoľko. U niektorých predstaviteľov sa so zvyšujúcim sa vekom bunky zvyšuje počet jadier. V jadre je vždy jedno jadierko.
Chloroplasty vegetatívnych buniek hnedých rias majú oko (stigma) a pyrenoidy, ktoré môžu byť jedno alebo viac; niektorým druhom chýbajú pyrenoidy. Ocellus, pozostávajúci z 30–40 lipidových guľôčok, funguje ako šošovka, ktorá sústreďuje svetlo na bičíkový opuch, ktorý je fotoreceptorom. Zoospóry a gaméty hnedých rias zvyčajne nemajú oko. V pyrenoidoch sa ukladajú náhradné látky. Chloroplasty sú najčastejšie malé, diskoidné, parietálne. Ich tvar môže byť hviezdicovitý, stuhovitý alebo lamelový. Tvar chloroplastov v bunkách hnedých rias sa môže meniť, keď bunka starne. Chloroplastový obal pozostáva zo štyroch membrán, pričom dve vonkajšie membrány sú spojené s EPS, endoplazmatickým retikulom bunky.
Celulóza, ktorá tvorí bunkovú stenu hnedých rias – niekedy nazývaná algullóza – sa svojimi vlastnosťami líši od celulózy vyšších rastlín. Mimo celulózovej steny je vysoko slizovitá pektínová vrstva, tvorená najmä kyselinou algínovou a jej soľami a zlúčeninami s bielkovinovými látkami. Kyselina algínová tiež sa vyskytuje iba v hnedých riasach.
Všeobecné charakteristiky.
Všetky hnedé riasy sú fototrofy. Produkty fotosyntézy - špecifický sacharid chrysolaminarín,alkohol manitol A lipidy. Lipidy sa hromadia vo forme kvapôčok oleja. Chrysolaminarín sa ukladá mimo chloroplastu: nachádza sa v špeciálnych vakuolách v cytoplazme v blízkosti pyrenoidu. Manitol vykonáva rezervu, ako aj osmoregulačnú funkciu; jeho koncentrácia vo vnútri bunky závisí od zmien slanosti vody.
Mladé bunky hnedých rias obsahujú malé a početné vakuoly, ktoré sa vekom v dôsledku fúzie zväčšujú. V cytoplazme existujú špeciálne formácie - fyzódy, ktoré obsahujú florotanínov. Tieto látky sú známe len u druhov hnedých rias. Funkcia florotanínov môže súvisieť s ochranou riasových stielok pred ich konzumáciou zvieratami, ako sú mäkkýše. Florotaníny hnedých rias pravdepodobne inhibujú usadzovanie epifytických rias a živočíchov na povrchu talu. Predpokladá sa, že florotaníny sa podieľajú na ochrane pred poškodením žiarením a na selektívnej absorpcii iónov ťažkých kovov z vodného prostredia. Bezfarebné florotaníny sa na vzduchu oxidujú za vzniku pigmentu fykofeínu, ktorý dáva sušeným hnedým riasam charakteristickú tmavú farbu.
Štruktúra talu. Medzi druhmi hnedých rias neexistujú jednobunkové a koloniálne formy, všetci jedinci sú mnohobunkové. Forma stielok hnedých rias je najrozmanitejšia. Sú to nite, plazivé alebo vertikálne stojace, jednoradové alebo viacradové; masívne pevné alebo kachľové kôry a kôry; tenké slizničné platničky alebo husté kožovité platničky na stonke, jednoduché alebo členité; bubliny a vrecká; Rúry a šnúry; rôzne rozvetvené tenké kríky alebo mohutné kríky. Pripevnenie tali sa vykonáva pomocou rizoidov alebo chodidiel. Na ich udržanie vo vertikálnej polohe vytvárajú veľké bentické formy hnedých rias vzduchové (plávajúce) bubliny naplnené plynom (obr. 29).
Ryža. 29. Vzhľad hnedých rias: 1 - Fucus; 2 - sacharín; 3 - Alaria; 4 - Sargasum
Thalli zástupcov radov Laminaria a Fucus sú najkomplexnejšie spomedzi hnedých rias. Majú znaky tkanivovej diferenciácie so špecializáciou buniek talu (obr. 30). V ich talí sa dá rozlíšiť štekať pozostávajúce z intenzívne zafarbených buniek a jadro pozostávajúce z bezfarebných buniek, ktoré sa často zhromažďujú vo vláknach. V Laminaria sa v jadre vytvárajú aj sitové rúrky a rúrkové vlákna. Jadro plní nielen transportnú funkciu, ale aj mechanickú, keďže obsahuje závity s hrubými pozdĺžnymi stenami. Medzi kôrou a jadrom mnohých hnedých rias môže byť medzivrstva veľkých bezfarebných buniek.
Výška stélka u predstaviteľov hnedých rias je najčastejšie interkalárna (interkalárna) a apikálna (apikálny), zriedkavo bazálny. Interkalárny rast môže byť difúzny alebo môže existovať zóna rastu. Vo veľkých formách sa interkalárny (interkalárny) meristém alebo aktívne deliaca skupina buniek nachádza v mieste prechodu "stopky" do "listovej" čepele. Veľké riasy majú na povrchu talu aj meristematickú zónu, takzvaný meristoderm (akási obdoba kambia vyšších rastlín).
reprodukcie u hnedých rias, vegetatívnych, asexuálnych a sexuálnych. Gaméty sa tvoria v mnohobunkových gametangiách, zoospórach - in 
Ryža. 30. Vnútorná štruktúra Laminaria(podľa: L.L. Velikanov a kol., 1991): A- pozdĺžny rez stonky; B- prierez stonky; IN- plátok taniera so sori zoosporangie
unilokulárna sporangia.
Vegetatívne rozmnožovanie v mnohých druhoch hnedých rias sa môže uskutočňovať na miestach stélky; v druhoch milý Sphacelaria- plodové vetvičky (plodové púčiky); u druhov rodu Fucus- skupina buniek na chodidle, ktoré sú schopné diferenciácie na nový slez.
K nepohlavnému rozmnožovaniu hnedých rias zvyčajne dochádza pomocou zoospór. Niektoré druhy sa rozmnožujú nepohyblivými tetraspórami alebo monospórami. Spóry nepohlavného rozmnožovania sa tvoria v dôsledku meiózy a následných mitóz v unilokulárna sporangia. Sexuálny proces je izogamia, heterogamia a oogamia. Gamety sa tvoria v mnohostennýtieto gametangie- špeciálne skupiny buniek. V každom „hniezde“ gametangia sa tvorí jedna gaméta.
Na hnedé riasy, sexuálne feromóny- rozpustné látky, ktoré koordinujú činnosť buniek pri rozmnožovaní. Feromóny, ktorých je známych najmenej desať, môžu buď stimulovať otváranie anterídií, alebo priťahovať mužské gaméty k ženským. Tiež sa podieľajú na izolácii druhov. Predpokladá sa, že feromóny sa tvoria vo väčšine druhov hnedých rias, ktoré majú sexuálny proces.
Životný cyklus. Hnedé riasy majú dva hlavné typy životných cyklov. Jeden typ je haplodiplobiont. Sporofyty sú diploidné, t.j. Všetky bunky obsahujú kompletnú sadu chromozómov. V unilokulárnych sporangiách sa tvoria spóry nepohlavného rozmnožovania. Pri tvorbe spór nastáva redukčné delenie – meióza. Takáto redukcia sa nazýva sporická. Bunky (spóry) vytvorené po meióze sú haploidné; obsahujú polovičný počet chromozómov. Haploidné zoospóry a tetraspóry vyklíčia do haploidného gametofytu, na ktorom sa tvoria gaméty vo viacbunkových gametangiách. Diploidná zygota vytvorená po splynutí gamét prerastie do diploidného sporofytu.
Ďalším typom je diplobiont; sporofyty, spóry a gametofyty sú diploidné; redukčné delenie nastáva pri tvorbe gamét. Takáto redukcia sa nazýva gametická. Po splynutí haploidných gamét vzniká diploidná zygota, ktorá vyklíči na diploidný sporofyt. Tento typ je bežný u druhov chalúh (obr. 31).

Ryža. 31. Životný cyklus Sacharíny(podľa: L.L. Velikanov et al., 1991): 1 – sporofyt, 2 – sori zoosporangií, 3 – zoospóry , 4 - gametofyt, 5 - anterídium, 6 - oogónium, 7 - anterozoid, 8 - zygota (oplodnené vajíčko)
Na makroskopickom sporofyte Sacharíny, v hornej časti dosky sa vytvárajú pruhy sori(skupiny) zoosporangií. V zoosporangiách sa po redukčnom delení vytvárajú zoospóry s dvoma nerovnakými bičíkmi. Rastú do mikroskopických gametofytov: samec s anterídiami a samica s oogóniou. Vajíčko oplodnené anterozoidmi okamžite, bez obdobia vegetačného pokoja, vyklíči a vytvorí sporofyt. V tomto životnom cykle Sacharíny končí.
Riasy Fucus nemajú sporofyty. O Fukusa mužské a ženské pohlavné orgány - anterídia so spermiami a plodnice s oogóniou a vajíčkami - sú umiestnené vo výklenkoch talu - scaphids, mužské a ženské scaphidia môžu byť na rovnakom nádoba alebo v rôznych receptoch. Oogonia sedí na nohe klietky , objavujúce sa na stene conceptcula(obr. 32, C, D). Bunka oogónia podlieha trom deleniam

Ryža. 32. Rozmnožovanie Fukusa(podľa: B. Fott, 1971; W. Braune a kol., 1974): A – fragment talu (1 - vzduchová bublina, 2 - recept); B– rez cez nádoby; IN– časť ženskej skafidie; G- oogónia; D- uvoľnenie vajec; E– časť samčieho scaphida; A– vetva s antheridia; 3 – výdaj spermií
jadier, prvým z nich je redukcia, v dôsledku čoho sa v oogóniu vytvorí 8 vajíčok.
Antherídia sa zvyčajne tvoria na rozvetvených vláknach, ktoré ich podporujú - parafýzy(obr. 32, E,A). V dôsledku niekoľkých delení sa vytvorí 64 spermií. spermie Fukusa majú oko umiestnené vo vnútri redukovaného chloroplastu. Na prednom konci spermie sa tvorí 13 mikrotubulov tulákprúd- špeciálna štruktúra, ktorá sa má podieľať na definícii ženského sexuálneho atraktantu.
Spermie sú priťahované k vajíčku Fukusa fukoserraten, nešpecifický feromón vylučovaný vajíčkom. Rozpoznanie na úrovni druhu je spojené so špecifickými oligosacharidmi na vajíčku a spermiách. Na povrchu vajíčka sú rôzne oblasti, z ktorých každá obsahuje špeciálne organické zlúčeniny - glykoproteíny. Spermie obsahujú glykoproteíny usporiadané do miest na membráne a výrastkoch predného bičíka, ako aj na povrchu samotnej spermie. Pri stretnutí s vajíčkom sa spermie pohybuje po škrupine vajíčka, akoby ho skúmala pomocou predného bičíka. Ak sú glykoproteíny spermií komplementárne ku glykoproteínom vajíčka, potom dochádza iba k rozpoznaniu spermie vajíčkom.
Systematika
Fosílne nálezy, ktoré môžu súvisieť s hnedými riasami, sú známe zo stredného silúru (asi 425 ma). Ale tieto nálezy sú tiež podobné niektorým moderným zeleným a červeným riasam. Fosílne nálezy, ktoré možno jednoznačne spájať s modernými hnedými riasami, pochádzajú z miocénu (5–25 miliónov rokov). Tieto riasy pripomína modernú Dictiotu. Vek hnedých rias stanovený molekulárnymi metódami je 155–200 miliónov rokov.
Na rozlíšenie radov hnedých rias sa používa typ organizácie talu, prítomnosť alebo neprítomnosť pyrenoidu, spôsob rastu, typ sexuálneho rozmnožovania (izogamia, heterogamia, oogamia) a životný cyklus. V posledných rokoch bol systém hnedých rias aktívne revidovaný. V rôznych klasifikačných schémach sa ako súčasť tried hnedých rias rozlišuje 7 až 14 rádov.
Desmarestiánsky poriadok–Desmarestiales. Riasy sú huňaté, viacnásobne rozvetvené, s terezovitým, plochým alebo splošteným talom. Pripevňuje sa k podkladu pomocou dobre vyvinutej podrážky. Na povrchu talu sa vyvíjajú chĺpky.
Rod Desmarestia (obr. 33). Hnedé alebo olivové kríky s dobre definovaným hlavným výhonkom, do výšky 0,8–1 m. Prichytené podrážkou v tvare kužeľa. Rozvetvenie 3-4 rádov, striedavé alebo opačné. Vetvy posledných rádov sú ploché alebo zriedkavejšie nitkovité. Druhy sú morské, žijú v arktických a miernych zemepisných šírkach v hĺbke 1–20 m, tvoria húštiny alebo rastú jednotlivo.

Ryža. 33. Desmarestia (r. Desmarestia)
Ralphsovský poriadok–Ralfsiales. Zástupcovia rádu majú talus vo forme kôr rôznych štruktúr alebo pozostávajú z bazálnej kôry a vertikálnych výhonkov.
Rod Analipus. Thallus sa skladá z trvalej kôrovitej bazálnej časti a z nej vybiehajú jednoročné olivovohnedé zvislé výhonky vysoké až 40 cm.Mnohobunkové výtrusnice sú valcovitého tvaru a dvojradovej štruktúry. Unilokulárne sporangie sú oválne a vyvíjajú sa medzi asimilačnými vetvami. Druhy rodu sú rozšírené v moriach miernych zemepisných šírok a tvoria pásy v prílivovej zóne.
Objednajte si Dictyota–Dictyotales.Pre druhy radu je charakteristický vrcholový rast. Vegetatívne bunky obsahujú početné diskovité chloroplasty bez pyrenoidov. Parenchymálny talus, charakteristické je izomorfné striedanie generácií. Pohlavný proces je oogamný, nepohlavné rozmnožovanie nepohyblivými spórami, spermie majú len jeden bičík. Bičík je dlhý, so sperenými výrastkami a smeruje dopredu. Zástupcovia rádu sa nachádzajú hlavne v tropických a menej často v miernych moriach.
Rod Dictyota (príloha, 4A). Má stélku vo forme plochých olivovohnedých, dichotomicky rozkonárených kríkov vysokých 11–15 cm.Vetvy sú lineárne, úzke, bez žiliek, s vidlicovitými vrcholmi. Tetrasporangia tvoria sori na povrchu talu, ktoré majú neurčitý alebo prstencový tvar. Distribuované v morských vodách miernych a teplých zemepisných šírok.
Objednajte si Laminaria–Laminariales. Laminaria majú stélku pozostávajúcu z dosky, stonky a upevňovacích orgánov - podošvy alebo rizoidov. Platnička je rovnomerná alebo zvrásnená, celá alebo členitá, s dierami, záhybmi, guľami alebo bez nich, od niekoľkých desiatok centimetrov do 20–40 m. Stonka je rozkonárená alebo častejšie nerozvetvená. Sporofyty sú jednoročné alebo viacročné, niektoré druhy majú 11–18 rokov. Slizničné kanály sa nachádzajú v kôre mnohých rias.
Najpopulárnejšími predstaviteľmi hnedých rias sú kelp (druhy rodov Laminaria alebo Saccharina), alebo „morské riasy“, ktoré sú rozšírené najmä v Tichom oceáne. Nachádzajú sa od hladiny (okraja) vody do hĺbky 200 m. Laminárie rastú prednostne v hĺbke 2–20 (30) m, na miestach s neustálym pohybom vody, prichytávané na kameňoch a skalách. Laminaria tvoria pásy rias pozdĺž pobrežia morí Ďalekého východu. Najsilnejšie húštiny sa nachádzajú v blízkosti Kurilských ostrovov a približne. Hokkaido, kde je sústredená asi polovica svetových zdrojov laminárií. Ročná svetová úroda chaluhových rias dosahuje 2 milióny ton čerstvej hmotnosti, pričom viac ako 1 milión ton pochádza z produkcie jej marikultúry v Číne.
Rod Sacharín (Laminaria). Thallus je dlhý od 0,5 do 5–10 m, má hrubú lesklú platňu, pružnú stonku a husto prepletené rizoidy, ktoré sú pripevnené k rôznym pevným substrátom. Okraje taniera sú hladké alebo zvlnené, na povrchu taniera sú často vydutia a záhyby. Na oboch stranách platničky sa tvoria nahromadenia sporangií - sori, po uvoľnení spór dochádza k deštrukcii vrchnej časti platničky. Žije od dvoch do niekoľkých rokov. Sacharíny sú cennou priemyselnou surovinou a predmetom marikultúry. Saccharina japonská tvorí húštiny v severnej polovici Japonského mora, ako aj v blízkosti južného a juhovýchodného pobrežia Sachalinu a južných Kurilských ostrovov. Saccharina japonská je najcennejším obchodným druhom, má vynikajúcu chuť a používa sa na prípravu rôznych jedál, konzerv, cukroviniek.
Rad Sphacelariae–Sphacelariales.
Hnedé riasy patriace do tohto rádu majú rozvetvený talus s vrcholovým rastom. Chlpaté konáre. Vegetatívne bunky majú početné chloroplasty bez pyrenoidov. Životný cyklus väčšiny s izomorfnou zmenou generácií, sexuálnym procesom je izogamia alebo heterogamia. Riasy tvoria malé hnedé trsy do výšky 3 cm a rozvetvené kríky do výšky 20 cm na kameňoch alebo iných riasach; existujú endofytickí zástupcovia.
Rod Sphacelaria. Má tenký talus. Vetviace sa vlákna tvoria malé, do 0,5–3 cm vysoké, hnedé trsy. Vegetatívne rozmnožovanie sa uskutočňuje špecializovanými, často trojlúčovými odnožami - propaguly. Sexuálny proces je izogamný. Ženské gaméty vylučujú feromóny ectocarpen a multifiden. Druhy rodu sú distribuované v moriach polárnych aj tropických zemepisných šírok.
Objednajte si Scytosiphon–Scytosiphonales. Zástupcovia s črevným, vezikulárnym, lamelárnym alebo vakovým, často dutým stélom s dĺžkou až 25 cm. Kôra je jednoradová alebo dvojradová, vnútorná časť (stena) talu je tvorená niekoľkými vrstvami veľkých buniek. Na povrchu talu sa vyvíjajú mnohobunkové sporangie, ktoré sa zhromažďujú v sori, ktoré podopierajú ich vlákna - parafýzy a chumáče chlpov.
Rod scytosifón je tenkostenná hnedá rúrka dlhá 4–25 cm (príloha 4B). Talus je pripevnený malou podrážkou a má zriedkavé priečne zúženia. Vyskytuje sa v pobrežnej zóne morí v rôznych zemepisných šírkach, často tvorí pásy v prílivovej zóne.
Fucus poriadok–Fucales. Majú trvalú stélku vo forme veľkých nepravidelne alebo dichotomicky sa rozvetvených kríkov do dĺžky 1–3 m. K podkladu sú pripevnené podrážkou v tvare kužeľa. Thallus má výhonky, fyloidy (listovité útvary), schránky (zásobníky gamét) a plávacie (vzduchové) mechúre. Nachádzajú sa v prílivovej zóne všetkých miernych a arktických morí, kde tvoria silné húštiny - pásy.
Rod sargasum(obr. 34, IN). Bohato rozvetvené olivovohnedé alebo hnedé kríky vysoké 0,5–1,5 m. Prichytené kužeľovou podrážkou. Fyloidy sa vyvíjajú na bočných výhonkoch vybiehajúcich z valcovej stonky v dolnej a strednej časti talu. Fyloidy sú kožovité, so stredovým rebrom, 0,2–2 cm dlhé. Plavecké mechúre sú oválne alebo zaoblené, s výrastkami alebo bez nich. Sargasy sú rozšírené v miernych a subtropických zemepisných šírkach od spodného litorálu do hĺbky 10 m, kde vytvárajú husté alebo riedke húštiny. V južnom Tichom oceáne – takzvanom Sargasovom mori – tvoria rozsiahle plávajúce akumulácie, v ktorých sa rozmnožujú len vegetatívne.
Rod Fucus (obr. 32).Thallus kožovitá, huňatá, konáruje sa dichotomicky, 25–50 cm vysoká, pri pobreží Ďalekého východu rastie niekoľko rokov, žije v prímorskej zóne. V plytkej vode často vytvára veľké húštiny. Druhy tohto rodu sa vyznačujú dichotomickými

Ryža. 34. Vzhľad riasy Fucus: A – Ascophyllum; B – Cystoseira; IN – sargasum; 1 - nádoba, 2 - plavecký mechúr
rozvetvený talus s plochými vetvami s pozdĺžnym rebrom, ktorý je pripevnený ku kameňom kužeľovou podrážkou. Fukusy sa používajú ako hnojivá, ako krmivo pre hospodárske zvieratá, na výrobu kŕmnej múky, alginátov, používajú sa v medicíne na liečbu strumy, korekciu hmotnosti atď.
Objednajte si ektokarpy–Ectocarpales. Riasy, ktoré sú tu zahrnuté, sú mikroskopické, voľne žijúce, epifyty, epizoity alebo endofyty. Reprodukčné orgány sú zastúpené unilokulárnymi (sporangia) a multilokulárnymi (gametangia) sporangiami; sú dlhovalcovité, krátkovalcovité alebo strukovité.
Rod ektokarpus. Thallus vo forme mäkkých hnedých kríkov do výšky 2 cm. Kríky sú tvorené jednoradovými, striedavo, dichotomicky alebo nesprávne rozvetvenými vláknami. Mnohobunkové výtrusnice sú strukovité, vytvorené na bočných konároch na 1–3-bunkovej stopke. Druhy sú rozšírené v moriach rôznych zemepisných šírok, v čistých a znečistených vodách, v litoráli a v hornej časti sublitorálu. Epifyty sa nachádzajú aj v znečistení antropogénnych substrátov.
Ekológia
Prevažná väčšina hnedých rias žije v morských vodách. V sladkých vodách sa vyskytuje iba 8 druhov. Existujú ročné aj viacročné druhy, ktorých vek môže dosiahnuť 15–18 rokov. Laminaria riasy Nereocystis A Macrocystis, ktoré žijú pozdĺž tichomorského pobrežia Severnej Ameriky, majú maximálnu veľkosť pre všetky riasy - až 30–40 m na dĺžku. Tvoria obrovské podmorské lesy v moriach Ďalekého východu. Hnedé riasy sa viažu na širokú škálu substrátov - kamene, kamene, štrk, lastúry a lastúry morských živočíchov a iné riasy. Niektoré malé formy hnedých rias žijú vo vnútri tkanív iných rias ako endofyty. Väčšina hnedých druhov žije v pripojenom stave. Tále odtrhnuté od zeme sú prúdom unášané do pokojných miest s bahnitým dnom a tam naďalej existujú. Druhy so vzduchovými bublinami na taluse, ktoré sa odtrhávajú od zeme, plávajú na povrch a vytvárajú veľké plávajúce zhluky, najmä v oblastiach so stabilným kruhovým prúdom, ako napríklad v Sargasovom mori. V takýchto zhlukoch existujú pomerne dlho a rozmnožujú sa len vegetatívne.
Hnedé riasy sú rozšírené vo všetkých moriach, no najväčší rozvoj dosahujú v moriach miernych a subpolárnych šírok. Ich veľké stélky rastú pripevnené na skalách a kameňoch a na tichých miestach blízko pobrežia a vo veľkých hĺbkach sa môžu dokonca držať na ventiloch lastúr mäkkýšov a štrku. Môžete ich stretnúť v rôznych hĺbkach – od prímorskej zóny, kde sú pri odlive celé hodiny mimo vody, až po 40–200 m. V oblasti Havajských ostrovov v hĺbke 180 m druhy rodu sargasum, A Kelp V Jadranské more bolo nájdené v hĺbke 200 m. Najhojnejšie húštiny hnedých rias sú pozorované v hĺbke 6–15 m, kde sú najlepšie svetelné podmienky a neustály pohyb vody, ktorý prináša živiny do ich slín a limitov usídlenie bylinožravých zvierat - fytofágy.
Trieda Zlaté riasy -Chrysophyceae
Jednobunkové, koloniálne alebo mnohobunkové organizmy, ktoré majú vo svojom životnom cykle zvyčajne bičíkové štádium. Bunky sú jednojadrové, s jedným alebo viacerými chloroplastmi. Zvyčajne je prítomná stigma (obr. 36). Bičíky 1–2, nerovnaké. Kontraktilná vakuola sa nachádza v prednej časti bunky. Vonkajšie obaly buniek týchto organizmov sú veľmi rôznorodé. U najprimitívnejších druhov je bunka odetá do jemného periplastu, ktorý umožňuje tvorbu výbežkov rôznych tvarov (rizopódia, pseudopódia), u iných je plazmalema pokrytá tvrdou celulózovou membránou, niekedy silne slizovou. Domy v zlatých riasach majú rôzne tvary: vázovité, guľovité, vajcovité, valcové; s jedným alebo viacerými otvormi. Klietka je pripevnená k základni domčeka ohybnou nohou alebo môže byť voľná. U niektorých chryzofytov sa na povrchu bunky ukladajú vápnité útvary – kokolity – rôznych tvarov a veľkostí a kremíkové bičíkovce majú vnútornú kremíkovú kostru. Rozmanitosť štruktúry týchto kostier umožňuje využiť pozostatky rôznych foriem fosílnych silikoflagelátov na určenie veku geologických hornín, ktoré ich obsahujú. U rozvinutejších predstaviteľov oddelenia zlatých rias sú bunky oblečené v škrupine pozostávajúcej z kremíkových šupín (niekedy nesúcich hroty) alebo sú uzavreté v domoch, cez otvory ktorých vychádzajú bičíky alebo pseudopódia.
Domy v druhoch zlatých rias sú vázovité, guľovité, vajcovité, valcovité, s jedným alebo viacerými otvormi. Klietka je pripevnená k základni domčeka ohybnou nohou alebo môže byť voľná.
Pigmenty - chlorofyly A A s fukoxantín. Chryzofyty sa vyznačujú najvyšším obsahom fukoxantínu spomedzi heterokontov (55 – 93 % všetkých karotenoidov).
Väčšina druhov sú sladkovodné fototrofy, ale nachádzajú sa aj heterotrofy a fagotrofy.
Náhradné látky - chrysolaminarín a oleje.
Medzi jednobunkovými formami zlatých rias prevládajú sladkovodné planktonické druhy a medzi mnohobunkovými formami bentické alebo epifytické. Nachádzajú sa hlavne v chladných morských a sladkých vodách. Je známych asi 360 druhov.

Ryža. 36. Zlaté riasy (podľa: C. Hoek van den et al., 1995): A- bunková štruktúra; B- radikulárny systém; IN- tripartitný mastigoném; G- rez časti chloroplastu; 1 - bazálne telo; 2 - bazálny opuch; 3 - krátky bičík; 4 - stigma; 5 - rizoplast; 6 - jadro; 7 - jadrová membrána; 8 - jadierko; 9 - mitochondrie; 10 - vakuoly; 11 - hlien; 12 - plazmalema; 13 - vezikuly; 14 - slizničné telo; 15 - lipidy; 16 - chloroplast; 17 - Golgiho prístroje; 18 - kontraktilná vakuola; 19 - dlhý bičík: 20 - 1. koreň; 21 - 2. chrbtica; 22 - 3. chrbtica; 23 - 4. koreň; 24 - chloroplastový nukleoid; 25 - HES; 26 - chloroplastová škrupina; 27 - pásová lamela; 28 - lamela; 29 - dlhé bočné vlákno; 30 - krátke bočné vlákno; 31 - koncové vlákno; 32 - tubulárna časť mastigonému
štruktúra talu u zlatých rias je najčastejšie monadická, ale môže byť veľmi rôznorodá: kokoidná, palmeloidná, plazmoidná, améboidná, filamentózna, parenchýmová (obr. 37).

Ryža. 37. Zlaté riasy (podľa: L.L. Velikanov et al., 1981): A – Ochromonas: 1 - vzhľad, 2 - cysta; B – Chromulina: 1 - vzhľad, 2 - cysta, 3 - schéma tvorby filmu z cýst; IN – Dinobrion: 1 - celkový pohľad na kolóniu, 2 - cysta; G – Chryzameba; D – gibberdia; E – Hydrurus: 1 - celkový pohľad na kolóniu, 2 - hrot vetvy, 3 - zoospóra
reprodukcie. V zlatých riasach existujú tri spôsoby reprodukcie: vegetatívna, asexuálna a sexuálna.
Vegetatívne rozmnožovanie sa vyskytuje pozdĺžnym delením buniek alebo rozpadom kolónie na časti v dôsledku delenia buniek v jednom, dvoch alebo troch smeroch.
sexuálny proces(bunky zlatých rias sú diploidné) - izogamia alebo konjugácia. V dôsledku sexuálneho procesu, ako aj za nepriaznivých podmienok, sa vytvárajú cysty s hustou škrupinou obsahujúcou kremík.
asexuálna reprodukcia realizované jedno- alebo dvojbičíkovými zoospórami, ktoré sa vyvíjajú v bežných vegetatívnych bunkách alebo zoosporangiách. Sexuálny proces: holo-, izo- a autogamia. Hologamia je typ sexuálneho procesu, pri ktorom sa dve bunky podobné vegetatívnym bunkám zlúčia na predných koncoch do dvojjadrovej zygoty, ktorá sa potom zmení na silicizovanú cystu.
Systematika
Zo zlatých rias bola koncom minulého storočia na základe výsledkov molekulárno-genetických štúdií izolovaná trieda Sinuraceae. Rozlišuje sa aj trieda Feotamniaceae - od žltozelených a zlatých rias. Preto sa rozsah triedy Chrysophyceae zmenšil av súčasnosti zahŕňa asi 360 druhov zaradených do troch rádov: Chromulinales, Hibberdiales a Hydrurales. Tvar, štruktúra a počet bičíkov, ako aj pigmentové zloženie a stavba tela sú dôležité systematické znaky.
Chromulínový poriadok - Chromulinales. Tento rád zahŕňa organizmy s monadickým, palmeloidným a améboidným typom diferenciácie talu. Monadické bunky s jedným bičíkom viditeľné pod svetelným mikroskopom.
Rod Chryzameba(obr. 37, A) - sladkovodné améboidné riasy, vo svojom životnom cykle majú bičíkové štádium. Zygota sa vyvinie do stomatocysty.
Rod Chromulina(obr. 37, IN) - jednobunkové voľne plávajúce organizmy, ktoré žijú hlavne v sladkých vodách. Bunky sú nahé; existuje jeden alebo dva zlaté plastidy.
Rod Dinobrion(Obr. 37 , G)- jednobunkoví a koloniálni, voľne plávajúci alebo pripútaní zástupcovia. Monády sú umiestnené vo vázovitých domčekoch tvorených celulózovými mikrofibrilami. Okrem celulózy obsahuje domček aj veľké množstvo aminokyselín. Huňatá forma kolónií je spojená s metódou vegetatívneho bunkového delenia, keď sa jedna z dcérskych buniek, opúšťajúca rodičovský dom, prichytí k jej otvoru. Otáčajúc sa okolo svojej osi tvorí svoj vlastný dom. Obe detské bunky môžu opustiť aj rodičovský dom. Bunky obsahujú 1-2 chloroplasty, na prednom konci bunky je viditeľné oko a 2 kontraktilné vakuoly. Chrysolaminarín sa nachádza vo vakuole na zadnom konci bunky. Reprodukcia je vegetatívna a sexuálna.
Objednajte si Gibberdiae–Hibberdiales. Zástupcovia tohto rádu sa vyznačujú jedinečným pigmentovým zložením. Okrem fukoxantínu majú dodatočný karotenoidový pigment, ktorý zachytáva svetlo, antheraxantín.
Zahŕňa pôrod Gibberdia, Chromophyton, Styloceras, Chrysopixis, Platyteka a ďalšie.
Rod gibberdia má dve štádiá životného cyklu: koloniálnu palmeloidnú nehybnú a jednobunkovú monadickú pohyblivú. Pri svetelnej mikroskopii je v monadickom štádiu viditeľný iba jeden bičík. Okrem fukoxantínu je charakteristická prítomnosť ďalšieho doplnkového svetlozberného karotenoidového pigmentu, antheraxantínu.
Objednajte si Hydrorus–Hydrurales. Zástupcovia tohto rádu majú talus s palmeloidnými a pseudoparenchymálnymi typmi štruktúry. Charakteristická je prítomnosť jedinečných tetraedrických zoospór. Nie je tam oko. Mitóza je polouzavretá.
Zahŕňa pôrod Hydrorus, Celloniella, Federmatium, Chrysonebula a ďalšie.
Rod Hydrurus(obr. 37, E) je medzi palmeloidnými riasami najviac diferencovaná. Jeho stélka sú veľké, až 30 cm dlhé slizké kolónie, ktoré vyzerajú ako hnedé rozvetvené šnúry, často vydávajú veľmi nepríjemný zápach. V kolóniách je možné rozlišovať medzi hlavným kmeňom a bočnými vetvami. Bunky ponorené do bežného hlienu obsahujú jeden miskovitý chromatofór a niekoľko kontraktilných vakuol. Bunky sú voľne umiestnené pozdĺž okraja kolónie a hustejšie v strede. Talus môže rásť len delením apikálnych buniek. Pri nepohlavnom rozmnožovaní sa v bunkách bočných vetiev kolónie tvoria zoospóry. Vytvára sférické cysty. Vyskytuje sa v horských potokoch a riekach so studenou vodou, kde sa prichytáva na tvrdých substrátoch.
Trieda rias Sinura -Sinurophyceae
syn- spolu a ura- chvost. Táto skupina organizmov bola izolovaná v roku 1987 zo zlatých rias. Zjednocuje monadické jednotlivé a koloniálne organizmy, niekedy so striedaním v životnom cykle monadického a palmeloidného štádia. Povrch pektínových škrupín je pokrytý škrupinou z kremíkových vločiek. Mitochondrie s tubulárnymi kristami sa zvyčajne nachádzajú v cytoplazme okolo chloroplastu. Jadro je jedno, chloroplasty obklopené štyrmi membránami, zvyčajne dvoma. Lamely sú tritylakoidné, existuje pásová lamela. Hlavnými pigmentmi sú chlorofyly A A s, β -karotén a fukoxantín. Neexistuje žiadna stigma. Bunky zvyčajne s 2 nerovnakými bičíkmi. Dlhý perovitý bičík smeruje dopredu. Krátky, hladký bičík, niekedy silne redukovaný, smerovaný dozadu.
Bunky sa rozmnožujú hlavne pozdĺžnym delením. Kolónie, ktoré sa rozpadajú, vedú k vzniku mladých kolónií. U niektorých druhov je sexuálny proces opísaný vo forme izogamie. Zároveň, na rozdiel od zlatých rias, k fúzii izogamét v sinurických riasach nedochádza na predných, ale na ich zadných koncoch. Silicifikované cysty s pórmi sa tvoria endogénne, podobne ako u chryzofytov.
Súčasťou bentosu sú prevažne planktónne formy, palmeloidné štádiá. Väčšina druhov rias sinura sú fototrofné sladkovodné organizmy.
Systematika.
Trieda rias Sinura zahŕňa 7 rádov: Chloramoebales, Synurales, Rhizochloridales, Ochromonadales, Heterogloeales, Parmales a Thallochrysidales.
Objednajte si Sinura - Synurales. Monadické formy, pektínové bunkové membrány sú zvyčajne pokryté plášťom kremičitých vločiek, stmelených organickou hmotou do jedného puzdra. Chloroplasty jeden alebo dva.
Rod Sinura(Obr. 38) - sladkovodné monadické formy s dvoma nerovnakými bičíkmi a dvoma parietálnymi chloroplastmi. Jadro je hruškovitého tvaru, nachádza sa v prednej časti bunky. Za jadrom je jedna veľká vakuola a v zadnej časti bunky je niekoľko malých kontraktilných vakuol. Bunky sú pokryté šupinami, ako dlaždice.Rozmnožovanie je najčastejšie vegetatívne, u niektorých je známy sexuálny proces. Vytvára kolónie (obr. 39, A). Často sa cysty objavujú vo všetkých bunkách kolónie. S masovým rozvojom Sinura môže spôsobiť nepríjemný zápach vody.

Ryža. 38. Vzhľad buniek Sinura
Rad Rhizochlorid - Rhizochloridales. Prevažujú sladkovodné organizmy s rhizopodiálnym typom diferenciácie talu, žijúce hlavne v sladkovodných útvaroch.
Rod Mallomonas(obr. 39, B-D) – jednobunková monáda s jedným dobre vyznačeným bičíkom a jedným rozdvojeným chloroplastom.

Ryža. 39. Vzhľad sinurických rias (podľa: G.A. Belyakova et al., 2006): A – štiepna kolónia Sinura; B, C – tvorba statospór a G – vzhľad Mallomonas
Bunka je pokrytá kachľovými a špirálovitými šupinami, niektoré majú ihlovité ostne (obr. 39, G). Charakteristická je prítomnosť statospór (obr. 39, B, C). U mnohých predstaviteľov je opísaný sexuálny proces, hologamia.
Rod Mixochlorisžije v bunkách listov sphagnum machov, má vzhľad veľkého mnohojadrového plazmódia. Na jeseň tvorí cysty, ktoré klíčia na jar. Vychádzajú z nich zoospóry alebo améby, ktoré prenikajú do prázdnych buniek listov a tam sa spájajú do plazmódia.
Rad Chloramoebaceae – Chloramoebales. Zahŕňa zástupcov monád. Nachádza sa v slaných a sladkých vodách.
Rod heterochloris – jeho bunky môžu meniť tvar a vytvárať pseudopódia. Tento jav je vlastný mnohým predstaviteľom rádu, ako aj tendencii k živočíšnemu spôsobu výživy. V bunke je niekoľko chloroplastov, olejových kvapiek a chrysolaminarínu. Pred bunkou sú kontraktilné vakuoly, v strede - jedno jadro. Rozmnožuje sa vegetatívne delením buniek.
Objednať Ochromonadales - Ochromonadales. Kombinuje holé formy s dvoma nerovnakými bičíkmi. Sladkovodné a morské formy.
Rod Ochromonas zahŕňa jednobunkové monády s dvoma nerovnakými bičíkmi. Bunky sú obalené iba plazmalemou. Na prednom konci bunky je kontraktilná vakuola a oko, na zadnom konci je vakuola s chrysolaminarínom. Rozmnožuje sa vegetatívne, štiepna brázda začína na prednom konci bunky medzi dvoma pármi bičíkových báz. Druhy rodu sú bežné v oligotrofných sladkých vodách, ale existujú aj morskí zástupcovia.
Rad Heterogleyes - Heterogloeales. Zahŕňa riasy s palmeloidným typom diferenciácie talu. Zástupcovia rádu sa vyskytujú častejšie v sladkých ako v slaných vodách.
Rod Helmintogleažije v brakických vodách. Táto kolónia sediaca na rozšírenej základni pozostáva z rozvetvených slizkých pásov. V tomto hliene sú náhodne usporiadané protoplasty, z ktorých každý je obklopený vlastným hlienovým obalom.
Trieda rias Phaeotamnia - Phaeothamniophyceae
Názov triedy pochádza z rodu typu Phaeothamnion(z gréčtiny. phaeos- hnedý ker). Zástupcovia tejto skupiny organizmov boli identifikovaní v oddelenej triede od žltozelených a zlatých rias v roku 1998 na základe analýzy sekvencie ribozomálnych génov a znakov bunkovej ultraštruktúry. Riasy Feotamnia sa vyznačujú unikátnou kombináciou pigmentov: fukoxantín s heteroxantínom, absencia violaxantínu.
Vlastnosti bunkovej štruktúry: absencia vakuol s chrysolaminarínom a absencia endogénnych cýst so stenami oxidu kremičitého.
Táto trieda zahŕňa jednobunkové, koloniálne a mnohobunkové organizmy s kokoidným, palmeloidným a vláknitým typom štruktúry talu. Bičíkovité štádiá s dvoma nerovnakými bičíkmi. Bičíky laterálne alebo subapikálne. Stigma sa nachádza v zoospórach. Pigmenty - chlorofyly A A s, β -karotén, fukoxantín, diadinoxantín, diatoxantín a heteroxantín. Hlavným náhradným produktom je paramylon ( β -glukán). Bunková stena je prítomná, pri delení buniek vzniká dcérska stena vnútri materskej steny.
V feotamniaceae, z hlavných spôsobov reprodukcie, sa nachádzajú iba vegetatívne a asexuálne. Nepohlavné rozmnožovanie sa uskutočňuje pomocou autospór alebo zoospór. Sexuálna reprodukcia nie je známa.
Druhy rias Feotamnia žijú výlučne v sladkých vodách. Systematika.
V súčasnosti je potvrdená monofyletická trieda rias feotamnia a medzi ochrofytmi má bližšie k hnedým a žltozeleným riasam ako k zlatým riasam. V súčasnosti je známych asi 30 druhov týchto rias, pripisovaných jednej rad Theotamnia - Phaeothamniales.
Rod Theotamnion predstavuje pripevnené vetviace nite do výšky 1 cm (obr. 40). Bunky sú valcovité, nepravidelné, rozšírené nahor, pozdĺž

Ryža. 40. Vzhľad Theotamnion.
dva až trikrát dlhšie ako široké. Počas delenia buniek sa materiál rodičovskej steny spotrebuje na vytvorenie vrstveného obalu obklopujúceho bunky vlákna. Na periférii bunky pod plazmatickou membránou sú vezikuly. Pripomínajú fyzódy – útvary nachádzajúce sa v bunkách hnedých rias. Bunky obsahujú jeden až niekoľko diskovitých olivovo-hnedých plastidov. Zoospóry sú tvorené 1–2, menej často 4–8 v jednej bunke. V zoospórach sú bičíky pripevnené bočne. Poloha bičíkových koreňov Theotamnion pripomína žltozelené a hnedé riasy. Chýba mu tiež rizoplast. V životnom cykle sa môžu vytvárať cysty, ale v ich stenách nie je kremík. Cysty klíčia ako zoospóry. Usadzuje sa ako epifyt na vláknitých riasach. Obýva stojaté a pomaly tečúce sladkovodné útvary.
Trieda Rafid rias -Raphidophyceae
Názov triedy pochádza z gréčtiny rhaphid - ihla. Zjednocuje jednobunkové biflagelátové organizmy bez bunkovej steny. Väčšinou sladkovodné organizmy so sploštenými bunkami (obr. 41).
Bunky majú bičíky nerovnakej dĺžky; bičík smerujúci dopredu je perovitý a dlhší; bičík smerujúci dozadu je hladký a kratší. Prechodná špirála chýba. Chloroplasty sú malé, početné alebo zriedkavejšie 1-2 na bunku, obklopené štyrmi membránami, z ktorých dve sú CES membrány. Tylakoidy sú usporiadané v hromadách po troch. Jadro je veľké, obklopené prstencom diktyozómov. Mitochondrie s tubulárnymi kristami. Existujú kontraktilné vakuoly. Cytoplazma je často vakuolizovaná. Bunky sú holé, obklopené iba plazmalemou. Stigma (oko) zvyčajne chýba.
Fotosyntetické pigmenty - chlorofyly A A s, β -karotén, vosheriaxantín, dinoxantín, diadinoxantín, heteroxantín.
Náhradnými asimilačnými produktmi sú tuky a oleje, menej často škrob a chrysolaminarín.
Žijú tiež v rašelinísk a iných biotopoch s kyslými a neutrálnymi reakciami vody. Niektoré druhy sa nachádzajú v brakických a morských vodách.

Ryža. 41. Vzhľad raphidových rias.
Systematika.
Veľmi dlho bola táto skupina rias vďaka svojej zelenej farbe, prítomnosti hltana a množstvu ďalších štruktúrnych znakov považovaná za triedu Chloromonas medzi euglenoidmi. Údaje z cytológie, biochémie, fyziológie a molekulárnej biológie však ukázali ich nepochybnú príslušnosť k oddeleniu rias Heterocont (Ochrophyte) a monofyletickú povahu skupiny rias Rafid.
Z jedinej je známych asi 25 druhov rad Huttonellaceae– Chattonellales.
Rod heterosigma(obr. 42, A) zahŕňa morské bičíkovce. Bunky oválneho tvaru s bičíkmi posunutými na stranu, plastidy sú umiestnené pozdĺž obvodu bunky. Neexistujú žiadne trichocysty. Plávajú a robia rotačné pohyby okolo pozdĺžnej osi tela. Môže spôsobiť toxické „kvety vody“ v pobrežných vodách.

Ryža. 42. Rafidové riasy (od: R. E. Lee, 1999): A – heterosigma; B – Goniostomum; 1 - chloroplast; 2 - mukocysta; 3 - kontraktilná vakuola; 4 - trichocysta; 5 - jadro
Rod Goniostomum(obr. 42, B) sú mobilné dorzoventrálne stlačené monády. Dorzálna strana je zakrivená, ventrálna strana je sploštená. Brázda na prednom konci vedie do trojuholníkového hltana, z ktorého vychádzajú dva dlhé bičíky; ich dĺžka je porovnateľná s dĺžkou bunky. Plastidy sa nachádzajú na okraji bunky. Pod plazmalemou sú trichocysty. Žije v sladkých vodách s kyslým pH.
Trieda rias Eustigma -Eustigmatophyceae
Názov triedy pochádza z gréčtiny EÚ- "dobre a stigma- "značka", "škvrna". Trieda spája nahé jednobunkové, menej často koloniálne organizmy prevažne kokoidnej štruktúry.
Jedno alebo viac jadier. Chloroplast je zvyčajne jeden, obklopený 4 membránami. Bičík je zvyčajne jedno predné perovité a druhé bazálne telo alebo zriedkavejšie dva nerovnaké bičíky. Stigma (oko) je prítomná, nachádza sa mimo chloroplastu, jeho granuly sú bez membrán.

Ryža. 43. Vzhľad rias Eustigma
Chloroplast je zvyčajne jednoduchý, veľký, viaclaločný, miskovitý alebo parietálny, žltozelenej farby. Tylakoidy sú zoskupené do lamiel v skupinách po troch, obopínajúca lamela chýba.
Hlavnými pigmentmi sú chlorofyl a, β-karotén, violaxantín a vosheriaxantín. Pyrenoid je zvyčajne prítomný iba vo vegetatívnych formách.
Náhradné látky sú oleje a látka neznámeho charakteru, ktorá sa ako pevný materiál ukladá mimo plastidov. Netvorí sa škrob.
Žijú v sladkých vodách, menej časté sú v moriach a v pôde.
Systematika.
Predtým bola táto skupina organizmov klasifikovaná ako žltozelené riasy. V súčasnosti sú riasy Eustigma považované za triedu v oddelení Heterocontophytes. Je známych asi 35 druhov, väčšinou sladkovodných, ktoré sa pripisujú jedinému Rad Eustigma - Eustigmatales.
Rod Eustigmatos(obr. 43) - jednobunkové drobné guľovité riasy. Bunková stena je hladká, celistvá, bez ornamentov. Žltozelený chloroplast jednoduchý, laločnatý, parietálny, so stopkovitými polyedrickými pyrenoidmi. Veľká centrálna vakuola s červeným obsahom. Rozmnožuje sa 2 alebo 4 autospórami alebo fľaškovitými zoospórami s jedným prichádzajúcim bičíkom, veľkou stigmou umiestnenou mimo chloroplastu na prednom konci a jedným zadným chloroplastom bez pyrenoidu. Bežná zložka pôdnej flóry. Známe z Nového Zélandu, Rakúska, Islandu az oblasti Arizony ako „horčičné morské riasy“.
Trieda Žltozelené riasy -Xanthophyceae
Žltozelené riasy sú tie, ktorých chloroplasty sú sfarbené do svetlej alebo tmavožltej, veľmi zriedkavo zelenej a len niekedy modrej farby. Farba stielky je určená prítomnosťou nasledujúcich pigmentov v bunkách chloroplastov - chlorofylov A A s, β -karotén a xantofyly. Prevaha posledného určuje originalitu farby žltozelených rias. Okrem toho paramylon, kvapká olej a len u niektorých druhov sa navyše v bunkách hromadia hrudky leukozínu a volutínu ako hlavný produkt asimilácie. Žltozelené riasy netvoria škrob. Charakteristickým znakom žltozelenej je prítomnosť monadickej štruktúry vo vegetatívnych bunkách a dvoch nerovnakých bičíkov v zoospórach. Bunková stena obsahuje celulózu, glukózu a urónové kyseliny. Bunková stena sa často skladá z dvoch častí.
Rozmnožovanie je vegetatívne, nepohlavné a sexuálne.
Široko distribuovaný v sladkých vodách. Zriedkavo sa vyskytuje v morských, brakických vodách a v pôde.
Predtým sa trieda žltozelených rias nazývala riasy Tribophytium, podľa typu rodu Tribonema (z gréčtiny. tribon – zručný, prefíkaný a nema – vlákno). Je známych asi 450 druhov.
Žlté zelené sa vyznačujú výraznou morfologickou rozmanitosťou. Medzi početnými predstaviteľmi tohto oddelenia sa nachádzajú takmer všetky hlavné typy stavby tela: améboidná, monadická, palmeloidná, kokoidná, vláknitá, multifilamentózna, lamelárna a sifonálna (obr. 44 – 46). Thallus jednobunkový,

Ryža. 44. Vzhľad žltozelených rias: 1, 2 - Charatiopsis, 3 – Centritractus, 4 – ophiocytium
koloniálne, mnohobunkové a nebunkové. Bunková membrána je hustá, pektín a celulóza, pozostávajúca z tesne sa prekrývajúcich častí alebo z dvoch ventilov. V škrupine je uložený oxid kremičitý alebo vápno. Väčšinou fixné formy. Medzi jednobunkovými druhmi existujú mobilné formy, ktoré nemajú hustú škrupinu a sú vybavené bičíkmi, lobopódiami a rizopódiami.

Ryža. 45. Vzhľad xantokokových žltozelených rias: 1–3 - Botrydiopsis, 4 – tetrahedriella, 5 – Pseudostaurastrum, 6 – Goniochloris, 7, 8 – Bumilleriopsis
Najviac žltozelené – nehybné organizmy. U mobilných jedincov sa pohyb môže vykonávať pomocou bičíkov alebo rizopódií. Bunky rôznych tvarov: guľovité, vretenovité, elipsoidné, valcovité, štvorstenné, kosáčikovité, hruškovité, vajcovité. Veľkosti Thallus od 0,5 – 1,5 µm ( Chloridella) do priemeru niekoľkých milimetrov ( Botrydiopsis) (obr. 45, 1 – 3) a dĺžky až do desiatok centimetrov ( Vosheria) (obr. 46, 3).

Ryža. 46. Vzhľad žltozelených rias: 1 - Tribonema, 2 – heteropédia, 3 – Vosheria, časť vlákna s oogóniom a anteridiom
Väčšina žltozelených druhov sú fototrofy, ale nachádza sa aj holozoické kŕmenie požitím baktérií a malých rias. Žltozelené riasy sú široko rozšírené v sladkých vodách. Časté sú aj v pôde, menej časté v morských a brakických vodách. Trieda zahŕňa aerobiontné, planktonické, bentické a perifytonické formy. Epifyty, epizoity, ako aj intracelulárne symbionty v bunkách prvokov.
Bez ohľadu na vonkajšiu štruktúru je vnútorná štruktúra bunky žltozelených rias rovnaká. V protoplaste sa zvyčajne nachádza niekoľko žltozelených chloroplastov, ktoré majú diskovitý, korytovitý, lamelárny, menej často stuhovitý, hviezdicovitý alebo miskovitý tvar s celistvými alebo laločnatými okrajmi. Sfarbenie je spôsobené absenciou fukoxantínu, ktorý je zodpovedný za zlatú a hnedú farbu iných ochrofytov. Z ostatných pigmentov majú β -karotén, vosheriaxantín, diatoxantín, diadinoxantín, heteroxantín. V mobilných formách sa červené oko alebo stigma zvyčajne nachádza na prednom konci chloroplastu. Len málo druhov má pyrenoidy poloponoreného typu. Jadro v bunke je zvyčajne jedno, má malú veľkosť, ale existujú druhy s viacjadrovými bunkami. Niektoré druhy majú jednu alebo dve kontraktilné (pulzujúce) vakuoly v prednej časti bunky.
Zástupcovia monád a pohyblivé štádiá (zoospóry a gaméty) majú dva nerovnaké bičíky. Výnimkou sú synzoospóry. Vosheri, v ktorej sa pozdĺž povrchu nachádzajú početné páry hladkých bičíkov mierne odlišných dĺžky. Krátky bičík končí akronémou. Bičíky sú pripojené k bunke subapikálne. Pri spermiách Vosheri bočné pripevnenie.
U druhov s améboidnou, monadálnou a palmeloidnou organizáciou bunková stena chýba, sú pokryté iba cytoplazmatickou membránou a môžu ľahko meniť tvar. Niekedy sa vo vnútri domov nachádzajú „holé“ bunky, ktorých steny môžu byť natreté hnedou soľou mangánu a železa. Prevažná väčšina foriem má bunkovú stenu pozostávajúcu z dvoch častí. V zložení bunkovej steny prevláda celulóza a obsahuje aj polysacharidy, pozostávajúce najmä z glukózy a urónových kyselín. V mladých bunkách je membrána tenká, s vekom sa zahusťuje. Môžu sa v ňom ukladať soli železa, ktorých zlúčeniny ho farbia do rôznych odtieňov hnedých a červených tónov. Častejšie je oxid kremičitý prítomný v bunkovej stene, čo jej dodáva tvrdosť a lesk. Môže byť tiež pokrytý vápnom a môže byť vytvarovaný rôznymi spôsobmi (tŕne, bunky, bradavice, zubáče, atď.) Výrastok škrupiny sa môže vytvoriť v pripojených formách – chodidlo s pripevňovacou podrážkou.
Vo filamentóznych formách žltozelených rias s lastúrnikmi sa pri rozpade filamentov bunkové membrány rozpadajú na úlomky v tvare písmena H. Tieto fragmenty sú pevne spojené polovice schránok dvoch susedných buniek (obr. 47). Počas rastu filamentov je fragment bunkovej steny dvoch susedných dcérskych buniek v tvare písmena H vložený medzi dve polovice membrány materskej bunky. Výsledkom je, že každá z dcérskych buniek je z polovice pokrytá starou membránou materskej bunky a z polovice – novovytvorená škrupina.

Ryža. 47. Schéma vzniku priečnej priehradky medzi dvoma dcérskymi bunkami vo vláknitých žltozelených riasach (podľa: A.A. Masyuk, 1993): A- fragment vlákna; B- položenie opaskového prstenca a vytvorenie priečnej priečky medzi dvoma bunkami; IN– vrstvenie membrán lastúrnikov; G– rozpad škrupiny na časti v tvare písmena H
V mobilných zástupcoch sú prítomné kontraktilné vakuoly. Zvyčajne sú 1-2 na bunku. Golgiho aparát zvláštnej štruktúry. Diktyozómy sú malé, obsahujú 3-7 cisterien. Jadro je jedno, menej často je veľa; u cenotických druhov sú bunky vždy viacjadrové.
reprodukcie. Väčšina druhov žltozelených rias sa vyznačuje vegetatívnym a nepohlavným rozmnožovaním.
Vegetatívne rozmnožovanie vykonávané rôznymi spôsobmi: bunkové delenie na polovicu, rozpad kolónií a mnohobunkových stielok na časti. O Vosheri vytvárajú sa špeciálne plodové puky.
O asexuálna reprodukcia Môžu sa tvoriť rôzne spóry: améboidy, zoospóry, synzoospóry, autospóry, hemizoospóry, hemiautospóry, aplanospóry. Zoospóry sú „nahé“ a zvyčajne majú hruškovitý tvar.
sexuálny proces- izogamia, heterogamia a oogamia - opísané u niekoľkých zástupcov. O Tribonemes gaméty majú podobnú veľkosť, ale líšia sa správaním - ide o izogamiu. O Vosheri pozoruje sa oogamia: na vláknach sa vytvárajú nádobky ženských gamét – oogónia a muž – anteridia.
Za nepriaznivých podmienok sa pozoruje tvorba cysty. Cysty (statospóry) sú endogénne, jednojadrové, zriedkavo viacjadrové. Ich stena často obsahuje oxid kremičitý a pozostáva z dvoch nerovnakých alebo zriedkavejšie rovnakých častí.
Systematika.
Na konci XIX – začiatkom 20. storočia do oddelenia zelených rias boli priradené rôzne rody žltozelených rias, čo súviselo predovšetkým s farebnou a morfologickou podobnosťou stél. Žlté zelené sa v súčasnosti považujú za triedu v rámci divízie ochrofytov.
Je známych asi 450 moderných druhov triedy žltozelených rias, ktoré sú zoskupené do štyroch rádov: Botridia, Mishococcus, Tribonem a Vosheria. Klasifikácia objednávok je založená na type diferenciácie talu a znakoch životného cyklu.
Rad Botridiae - Botrydiales. Rad zahŕňa druhy so sifonálnym typom diferenciácie talu, v ktorých nie je žiadny oogamný pohlavný proces.
Rod Botridiumžije na pôde a má vzhľad zelených bublín s veľkosťou niekoľkých milimetrov, pripevnených bezfarebnými rizoidmi. Talus je sifonálny, obsahuje početné jadrá a plastidy. Škrupina je viacvrstvová, môže sa na ňu ukladať vápno. Rozmnožovanie je nepohlavné pomocou biflagelátových zoospór, pričom celý obsah močového mechúra sa rozpadá na jednojadrové úlomky. Pri nedostatku vlahy sa rozmnožuje pomocou aplanospór alebo tvorí hrubostenné cysty. V niektorých prípadoch celý obsah močového mechúra ide do tvorby jednej veľkej cysty. V iných prípadoch sa cysty tvoria v rizoidoch, kde najskôr prechádza obsah močového mechúra. Cysty klíčia buď priamo do nového talu, alebo tvoria zoospóry. Sexuálny proces je izo- a heterogamia. Zygota klíči okamžite, bez obdobia pokoja. Bežné a rozšírené druhy v suchozemských biotopoch, vyskytujúce sa pozdĺž brehov potokov, rybníkov alebo na pôdach bez vegetácie.
Objednajte si Mischococcal–Mishococcales. Jednobunkoví alebo koloniálni zástupcovia s kokoidným typom diferenciácie talu.
Rod Charatiopsis zahŕňa jednobunkové pripojené formy. Pri rozmnožovaní vytvára zoospóry, aplanospóry a hrubostenné cysty (obr. 44, 1-2).
Rod ophiocytium(obr. 44, 4) má podlhovasté valcové články, ktoré môžu byť rovné, ohnuté alebo špirálovito stočené a na konci môžu niesť bodec. Bunková stena sa skladá z dvoch nerovnakých častí, z ktorých väčšia časť sa podieľa na raste buniek, menšia časť je trvalá a má tvar čiapočky. Jednobunkové a koloniálne druhy, voľne žijúce alebo pripevnené k substrátu malou stopkou. Rozmnožujú sa zoospórami a aplanospórami, nachádzajú sa cysty. Žijú v sladkých vodách.
Rod Mischococcus vytvára stromovité prirastené kolónie. Vetvenie je dichotomické a tetrachotomické. Bunky sú umiestnené v 2 alebo 4 na vrcholoch hlienových vetiev kolónie. Bunky sú guľovité až oválne, s tenkou alebo hrubou bunkovou stenou. Niekedy je bunková stena lesklá a hnedá kvôli jej impregnácii soľami železa. Mladé jednobunkové organizmy so slizkou diskovitou základňou, ktorá slúži ako opora na pripevnenie. Po uvoľnení spór sa protoplast materskej bunky zmení na rôsol a natiahne sa, dĺžka sa zväčší 6-krát ako šírka, a tak sa objaví valcovitá noha. Základom stonky sa vždy stáva prázdna bunková stena materskej bunky. Nepohlavné rozmnožovanie zoospórami a autospórami. Autospóry sú pripevnené k hornému okraju hlienovej stopky. Následné bunkové delenia opakujú proces a vytvárajú kolóniu podobnú stromu. sexuálny proces – izogamia. Žijú v malých sladkovodných útvaroch ako epifyty vláknitých rias. Známy v strednej Európe a Ázii.
Rad Tribonemaceae - Tribonematales. Zástupcovia majú vláknitý, multifilamentózny, falošný tkanivový a tkanivový typ diferenciácie talu. Bunkové steny sú buď s prekrývajúcimi sa časťami v tvare H alebo sú plné.
Rod Tribonema- nerozvetvené nite (obr. 46, 1). Bunky sú valcovité alebo súdkovité. Bunková stena sa skladá z dvoch polovíc, ktoré sa nachádzajú s okrajmi na sebe v strede bunky. Mušle sú často vrstvené. Úlomky nití vždy končia prázdnymi polovicami úlomkov mušlí v tvare písmena H, ktoré vyzerajú ako vidlica. V bunkách je niekoľko žltozelených plastidov, nie sú tam žiadne pyrenoidy. Rozmnožovanie je vegetatívne (fragmentácia filamentov), asexuálne (zoospórami a aplanospórami) a sexuálne (izogamia), pričom aplanospóry sú produkované častejšie ako zoospóry. Môže vytvárať akinéty. Žijú v sladkých vodách, kde sa obzvlášť hojne rozvíjajú v chladnom období.
Objednajte si Vosherievye - Vaucheriales. Všetci zástupcovia majú sifonálny talus, oogamný sexuálny proces a synzoospóry.
Rod Vosheria(obr. 46, 3) má talus nebunkovej štruktúry; jeho stélka dosahuje dĺžku niekoľko centimetrov, prichytáva sa k substrátu pomocou bezfarebného rizoidu. V závitoch nie sú žiadne priečky, väčšina talu je obsadená vakuolou a početné jadrá a plastidy sa nachádzajú pozdĺž periférie v cytoplazme. Nite s vrcholovým rastom a zriedkavým bočným vetvením. Priečky sa vytvárajú pri poškodení talu a na oddelenie reprodukčných orgánov. Nepohlavné rozmnožovanie sa uskutočňuje pomocou aplanospór, synzoospór, akinét. Synzoospóry sa tvoria po jednej v zoosporangiu, ktoré je oddelené od vegetatívnych buniek priehradkou na konci vlákna. Zoospóry sú viacjadrové a mnohobičíkové. Sexuálny proces je oogamia. Zygota je pokrytá hrubou škrupinou a po období pokoja vyrastie do nového talu.
Druhy Vosheriširoko rozšírený v sladkých, brakických a morských vodách, ako aj v suchozemských biotopoch. Vyskytujú sa na všetkých kontinentoch vrátane Antarktídy. Tvoria trávovozelené alebo tmavozelené prepletené hmoty - takzvané rohože, hladké, plazivé alebo vankúšovité. Vodné, polovodné, suchozemské formy. Žijú v rôznych biotopoch: moria, ústia riek, ústia riek, slané močiare, mangrovy, potoky, kanály, jazerá, rybníky, orná pôda a močiare.
Významheterokontné riasy
Z oddelenia rias Heterocont majú pre prírodné ekosystémy a pre človeka najväčší význam hnedé riasy.
Hnedé riasy - hlavné zdroj organickej hmoty v pobrežnej zóne morí. Ich biomasa v moriach mierneho a subpolárneho pásma môže dosahovať niekoľko desiatok kilogramov na meter štvorcový. Húštiny hnedých rias poskytujú podmienky na kŕmenie a rozmnožovanie mnohých pobrežných živočíchov a iných rias. Charles Darwin pozoroval hnedé riasy pri pobreží Južnej Ameriky Macrocystis, napísal: „Tieto rozsiahle podvodné lesy južnej pologule môžem porovnať iba s pozemskými lesmi tropických oblastí. A predsa, ak by sa v niektorej krajine zničil les, nemyslím si, že by zahynul ani približne rovnaký počet živočíšnych druhov ako pri ničení tejto riasy.
Poslúžia húštiny hnedých rias miesto na jedlo,prístrešie a chov veľa zvierat. Obrazne povedané, hnedé riasy poskytujú iným vodným organizmom „stôl, úkryt a škôlku“.
Hnedé riasy sú široko používané ľuďmi. Sú bohatí jód a ďalšie mikroživiny. Národy juhovýchodnej Ázie ich tradične používajú pre jedlo, najmä zástupcovia rádu kelu, z ktorých sa pripravuje široká škála jedál. Kŕmiť múku, pripravený z hnedých rias, zvyšuje produktivitu hospodárskych zvierat; zároveň sa vo vajciach a mlieku zvyšuje obsah jódu.
Z hnedých rias prijímaťalgináty- soli kyseliny algínovej. Algináty sú široko používané v rôznych priemyselných odvetviach. Ide o netoxické zlúčeniny s koloidnými vlastnosťami, preto majú široké využitie v potravinárskom a farmaceutickom priemysle. Kyselina algínová a jej soli sú schopné 200-300-násobne absorbovať vodu a vytvárať gély, ktoré sa vyznačujú vysokou odolnosťou voči kyselinám. V potravinárstve sa používajú najmä ako emulgátory, stabilizátory, želírujúce a vodu zadržiavajúce zložky. Napríklad suchý práškový alginát sodný sa používa pri výrobe práškových a briketovaných rozpustných produktov (káva, čaj, sušené mlieko, želé atď.) na ich rýchle rozpúšťanie. Vodné roztoky alginátov sa používajú na mrazenie mäsa a rybích výrobkov. Až 30 % z celkového objemu alginátov vyrobených vo svete smeruje do potravinárskeho priemyslu.
V textilnom a celulózo-papierenskom priemysle sa algináty používajú na zahusťovanie farieb a zvýšenie pevnosti ich väzby s podkladom. Impregnácia látok niektorými alginátmi im dodáva ochranné vlastnosti: vodeodolnosť, odolnosť voči kyselinám a zvyšuje mechanickú pevnosť. Na získanie umelého hodvábu sa používa množstvo solí algínových kyselín. Počas druhej svetovej vojny v USA a Anglicku sa z kyseliny algínovej a jej solí vyrábalo veľké množstvo maskovacích látok a sietí pre obytné a priemyselné budovy.
Algináty sa používajú v metalurgii: v zlievárňach zlepšujú kvalitu formovacej zeminy. Pri výrobe elektród na elektrické zváranie sa používajú soli kyseliny algínovej, ktoré umožňujú získať kvalitnejšie zvary. Algináty sa používajú aj pri výrobe plastov, syntetických vlákien, náterov a stavebných materiálov odolných voči poveternostným vplyvom. Používajú sa pri výrobe vysokokvalitných mazív pre stroje. V rádiovej elektronike pôsobia algináty ako spojivo pri výrobe vysokokvalitných feritov.
Najväčšie uplatnenie nachádza vo vode rozpustný alginát sodný, schopný vytvárať viskózne roztoky. Je široko používaný na stabilizáciu rôznych roztokov a suspenzií. Pridanie malého množstva alginátu sodného do potravinárskych výrobkov - konzervy, zmrzlina - zlepšuje ich kvalitu. Vyrába sa z neho aj dekoratívna kozmetika, krémy a masky v parfumérskom priemysle.
Vo farmaceutickom priemysle sa algináty používajú na poťahovanie tabliet, piluliek, ako základ pre rôzne masti a pasty, ako gély na nosiče liečiv a pri výrobe rozpustných chirurgických stehov. V medicíne sa alginát vápenatý používa ako hemostatické činidlo a ako sorbent, ktorý odstraňuje rádionuklidy (napríklad stroncium). Ročná produkcia alginátov vo svete presahuje 20 tisíc ton.
Ďalšou dôležitou látkou získanou z hnedých rias je šesťsýtny alkohol manitol. Manitol sa používa ako náhrada cukru pre diabetikov. Okrem toho sa môže použiť ako náhrada plazmy na konzerváciu krvi. Používa sa na výrobu tabliet vo farmaceutickom priemysle. Manitol sa tiež používa v výroba syntetických živíc, farieb, papiera, výbušnín, apretúry kože.
fukoidany, získané z hnedých rias sú účinné antikoagulanciá, dokonca aktívnejšie ako heparín. Ich použitie na výrobu protinádorových liečiv a antivírusových zlúčenín sa považuje za sľubné. Koniec koncov, aj pri najnižších koncentráciách môžu fukoidany inhibovať prichytenie vírusov na povrch buniek. Fukoidany majú tiež schopnosť tvoriť mimoriadne silný a viskózny hlien, ktorý sa využíva pri príprave stabilných emulzií a suspenzií.
Energetická kríza, ktorá v posledných rokoch zachvátila mnohé krajiny sveta, viedla k potrebe hľadania nových netradičných zdrojov energie. Takže v USA sa na tento účel skúma možnosť šľachtenia obrovských rias. Macrocystis s následným spracovaním na metán. Vypočítalo sa, že z plochy 400 km 2, ktorú zaberá táto riasa, možno získať 620 miliónov m 3 metánu.
Heterokontné riasy z tried Golden, Yellow-green, Sinura, Rafidophyte a Eustigma riasy, zastúpené najmä mikroskopickými organizmami, sú rozšírené v sladkovodných útvaroch všetkých klimatických pásiem zemegule, ale sú bežnejšie v miernych zemepisných šírkach. Medzi zlatými riasami sú druhy, ktoré žijú v moriach a slaných jazerách a veľmi málo v znečistených vodách. Zlaté riasy dosahujú maximálny rozvoj v chladnom období: v planktóne dominujú skoro na jar, neskoro na jeseň av zime. V tejto dobe zohrávajú významnú úlohu ako producenti prvovýroby a slúžia ako potrava pre organizmy zooplanktónu.
Napríklad nejaké zlaté riasy Uroglena A Dinobrion, vyvíjajú sa vo veľkom množstve, sú schopné spôsobiť vodné kvitnutie. Uvoľňujú aldehydy a ketóny, ktoré môžu dodávať vode nepríjemný zápach a chuť, a Uroglena– mastné kyseliny toxické pre ryby.
Rafidové riasy sú široko zastúpené v planktóne sladkovodných útvarov s prevažne kyslým pH, najmä v rašelinísk, menej často vo veľkých jazerách. V sladkovodných útvaroch sa môžu vytvárať miestne „kvety“. Goniostomum. Rafidové riasy sa nachádzajú aj v odsolených morských zálivoch a kalužiach na pobreží, ako aj na otvorenom mori. S masovým rozvojom v pobrežných morských vodách spôsobujú toxické "kvety" vody. Takže pri pobreží Kanady počas „kvitnutia“ môže koncentrácia buniek rapídnej riasy Heterosigma dosiahnuť 30 miliónov na 1 liter. Ohniská raphidových rias často vedú k rozvoju „červených prílivov“, ktoré sú spojené s úhynom rýb. Príčinou takýchto "červených prílivov" môžu byť druhy pôrodu Hattonella, Olistodiscus, Heterosigma a Fibrocapsa.
Sinura riasy s hromadným vývojom v sladkovodných útvaroch môžu vode spôsobiť nepríjemný zápach ( Sinura). Riasy Feotamnia sa nachádzajú v stojatých a pomaly tečúcich sladkých vodách, kde sa epifyticky usadzujú na vláknitých riasach.
Riasy Eustigma sa nachádzajú iba v sladkej vode alebo v pôde.
Žltozelené riasy sú rozšírené na všetkých kontinentoch, žijú najmä v sladkej vode a pôde, ako aj v suchozemských, brakických vodách a morských biotopoch. Žltozelené riasy obývajú čisté a znečistené vody s rôznymi hodnotami pH: môžu žiť v kyslých aj zásaditých vodách. Vyskytujú sa hlavne v čistých sladkovodných nádržiach, menej často v moriach a brakických vodách, preferujú mierne teploty, častejšie sa rozvíjajú na jar a na jeseň, hoci existujú druhy, ktoré sa vyskytujú počas všetkých ročných období vrátane zimy. Najčastejšie ich možno nájsť v zhlukoch nití a medzi húštinami vyšších vodných rastlín v pobrežnej zóne riek, rybníkov, jazier a nádrží.
Prevažná väčšina žltozelených sú voľne žijúce formy, ale v bunkách prvokov sú aj vnútrobunkové symbionty – zooxantely. Zaujímavú vnútrobunkovú symbiózu tvoria chloroplasty morských druhov Vosheri s lastúrnikom Elision. Počas deviatich mesiacov je tento mäkkýš schopný fotoautotrofnej fixácie oxidu uhličitého v kultúre. Ide o najdlhšiu symbiózu tohto typu, kedy je symbiotický plastid v priamom kontakte s cytoplazmou zvieraťa. V prírode sa larvy mäkkýšov živia vláknami. Vosheri. V dôsledku fagocytózy sa chloroplasty rias dostávajú do cytoplazmy epitelových buniek mäkkýšov. Počas tohto procesu sa chloroplastová membrána stáva trojvrstvovou, jedna vonkajšia membrána CES (chloroplastové endoplazmatické retikulum) sa stráca. Tento jav je dobrým dôkazom toho, že v priebehu evolúcie by mohli vzniknúť chloroplasty s tromi membránami ako výsledok sekundárnej symbiogenézy v dôsledku straty membrán.
Žltozelené, zlaté a iné heterokontné riasy sú producentmi kyslíka a organických látok; sú súčasťou potravinového reťazca. Heterokontofyty sa podieľajú na samočistení znečistených vôd a pôd, tvorbe sapropelu, na procese akumulácie organickej hmoty v pôde, ovplyvňujúcej jej úrodnosť. Používajú sa ako reprezentatívne organizmy pri určovaní stavu znečistenia vôd; žltozelené riasy sú súčasťou komplexu mikroorganizmov používaných na čistenie odpadových vôd.
Kontrolné otázky
Vymenujte charakteristické znaky štruktúry hnedých rias.
Štrukturálne znaky hnedých rias thalli.
Ako sa rozmnožujú hnedé riasy? Čo sú monospóry, tetraspóry a zoospóry, izogamia, heterogamia a oogamia?
Aké sú životné cykly hnedých rias? Reprodukcia fucus a chaluhových rias.
Vymenujte charakteristické znaky a typických predstaviteľov radov hnedých rias.
V ktorých biotopoch sa nachádzajú hnedé riasy? Aký je ich význam v prírode.
Hospodársky význam hnedých rias.
Vymenujte charakteristické znaky stavby a typických predstaviteľov zlatých rias.
Aké pigmenty a druhy výživy sú známe u zlatých rias?
Rozmnožovanie a ekológia zlatých rias.
Vymenujte charakteristické znaky a typických predstaviteľov žltozelených rias.
Aké pigmenty a druhy výživy sú známe u žltozelených rias?
Ako sa rozmnožujú žltozelené? Typy sexuálneho rozmnožovania: izogamia, heterogamia a oogamia?
Vymenujte charakteristické znaky stavby a typických predstaviteľov sinurických rias.
Rozmnožovanie a ekológia sinurických rias.
Vymenujte charakteristické znaky stavby a typických predstaviteľov rias feotamnia.
Rozmnožovanie a ekológia rias feotamnia.
Vymenujte všeobecné znaky stavby a typických predstaviteľov rafidofytných rias.
Rozmnožovanie a ekológia rafidofytných rias.
Aké sú spoločné znaky štruktúry a typických predstaviteľov rias Eustigma.
Rozmnožovanie a ekológia rias Eustigma.
Hodnota heterokontných rias v prírodných ekosystémoch.
Bičíkovce - najstaršia skupina prvokov, ležiaca pri koreni genealogického stromu živočíšneho sveta a spájajúca živočíchy so svetom rastlín. Ešte v polovici predminulého storočia známy ruský biológ L. S. Tsenkovsky (1822-1887) poukázal na absenciu ostrej hranice medzi bičíkovitými prvokmi a jednobunkovými riasami. Niektoré bičíkovce (euglenoidy a fytomonády) sú totiž oprávnene klasifikované ako druh prvokov, t.j. živočíchov, ako aj typ zelených rias, t.j. rastliny.
Charakteristika triedy
Jedným z najdôležitejších znakov používaných pri klasifikácii prvokov sú organely pohybu. U bičíkovcov sú to zväzky alebo metličky - tenké cytoplazmatické výrastky. Zvyčajne sa nachádzajú na prednom konci tela a vznikajú zo špeciálnych bazálnych zŕn (bazálneho telieska alebo kinetozómu). Počet bičíkov u rôznych druhov sa pohybuje od 1 do 8 alebo viac. V niektorých bičíkoch (leishmánia, trypanozómy) je na báze bičíka navyše umiestnený špeciálny organoid, kinetoplast. Vo svojej ultraštruktúre zodpovedá mitochondriám, ale vyznačuje sa vysokým obsahom DNA. Predpokladá sa, že v kinetoplaste sa generuje energia na pohyb bičíka, ktorý vykonáva rotačný pohyb a akoby sa zaskrutkoval do vody. U niektorých predstaviteľov triedy bičík prebieha pozdĺž tela a spája sa s ním tenkým výrastkom cytoplazmy. Uvedený výrastok alebo zvlnená membrána vykonáva vlnité pohyby a slúži ako dodatočná organela pohybu.
Podľa spôsobu výživy (asimilácie) sa delia na:
Rozmnožovanie je zvyčajne nepohlavné, pozdĺžnym delením na dve časti. Mnohé druhy majú aj pohlavný proces, počas ktorého sa sexuálne formy spájajú (kopulácia).
V stojatých sladkovodných nádržiach žijú početné druhy zelených bichenos (euglena a fytomonády). Občas sa objavia v takom veľkom množstve, že voda zozelenie (vodný kvet).
Samotárske voľne žijúce formy
Typickým predstaviteľom jednotlivých foriem je orlíček zelený (Euglena viridis). Jeho tvar tela je vretenovitý, konštantný v dôsledku zhutnenia vonkajšej vrstvy protoplazmy. V cytoplazme, v zadnej časti tela, je veľké sférické jadro.
Na prednom konci tela zeleného euglena je tenký bičík, v dôsledku ktorého sa pohybuje. Nachádza sa tu aj pulzujúca vakuola, jej rezervoár a niekoľko chromatofórov obsahujúcich chlorofyl, v ktorých sa na svetle syntetizujú sacharidy z oxidu uhličitého a vody (fotosyntéza), t.j. pozoruje sa autotrofná výživa. Produkty fotosyntézy sa ukladajú vo forme látky podobnej škrobu – paramylu. Euglena hľadá osvetlené miesta pomocou svetlocitlivého oka – do červena sfarbeného fotoreceptora, umiestneného tiež na prednom konci tela.
Euglena sa môže živiť aj holozoickým spôsobom, prehĺtajúc tvarované častice potravy. Ak je prostredie bohaté na rozp organickej hmoty, potom vstupujú do tela euglena osmózou. V štruktúre a spôsobe výživy euglena je teda zaznamenaná kombinácia vlastností živočíšneho a rastlinného organizmu.
Udržiavanie určitej koncentrácie solí v cytoplazme, t.j. osmotického tlaku, sa vykonáva špeciálnym organoidom - pulzujúcou alebo kontraktilnou vakuolou umiestnenou na prednom konci tela. Pulzujúca vakuola periodicky zväčšuje svoj objem. Po dosiahnutí určitej veľkosti vyleje nahromadenú kvapalinu cez nádrž a po chvíli začne znova rásť. Kvapalina vstupuje do vakuoly cez adduktorové kanály.
Rozmnožovanie Euglena je nepohlavné, pozdĺžnym delením tela. Najprv sa rozdelí jadro, zdvojnásobí sa bazálne telo, chromatofóry a potom sa rozdelí cytoplazma. Bičík sa odhodí alebo prejde na jedného jedinca a u iného sa vytvorí nanovo. Delenie v pozdĺžnom smere je jedným z charakteristické znaky trieda bičíkov. Za nepriaznivých podmienok nastáva encystácia: bičík sa stiahne, euglena sa zaobľuje a vytvára hustú schránku. Encyklovaná euglena sa môže rozdeliť jeden alebo viackrát, zatiaľ čo zostáva vo vnútri škrupín.
Rôzne typy euglena sú charakteristické pre vodné útvary s rôznym stupňom organického znečistenia, preto spolu s riasami môžu slúžiť ako kritérium pre hygienické hodnotenie zdrojov zásobovania vodou.
Koloniálne formy

Koloniálne bičíkovce (volvox, pandorina, eudorina atď.) sú považované za prechodné formy od jednobunkových k mnohobunkovým organizmom. Najjednoduchšie usporiadané kolónie pozostávajú zo 4-16 úplne identických jednobunkových jedincov spojených dohromady - zooidov. Každý zooid má bičík, ocellus, chromatofóry a kontraktilnú vakuolu.
Zástupca koloniálneho bičíkatého druhu - volvox (Volvox globator) tvorí veľké guľovité kolónie pozostávajúce z mnohých tisícok vegetatívnych zooidov - malých buniek hruškovitého tvaru, z ktorých každá má dva bičíky. Priemer gule je 1-2 mm. Jeho dutina je vyplnená želatínovou hmotou. Všetky bunky Volvox (zooidy) sú navzájom prepojené tenkými protoplazmatickými mostíkmi, čo umožňuje koordinovať pohyb bičíkov. Kolónia sa vo vode pohybuje vďaka koordinovanému pohybu bičíkov jednotlivých jedincov.
Vo Volvoxe je už pozorované rozdelenie funkcie buniek kolónie. Takže na jednom póle kolónie, ktorým sa pohybuje dopredu, sú bunky s vyvinutejšími očami citlivými na svetlo a v spodnej časti kolónie (kde sú oči slabo vyvinuté) sú bunky schopné delenia (reprodukčné bunky, generatívne zooidy), t.j. je zaznamenaná diferenciácia na somatické a sexuálne jedince.
Reprodukcia Volvoxu sa uskutočňuje na úkor špeciálnych - generatívnych - zooidov. Opúšťajú povrch vo vnútri kolónií a tu, množiac sa delením, vytvárajú dcérske kolónie. Po smrti materskej kolónie začínajú dcéry samostatný život. Na jeseň sa vplyvom generatívnych jedincov vytvárajú aj sexuálne formy: veľké nepohyblivé makrogaméty (samičie reprodukčné zooidy) a malé mikrogaméty vybavené dvoma šnúrami (samčie reprodukčné zooidy). V procese gametogenézy sa jednotlivci, ktorí sa zmenia na makrogaméty, nerozdelia a nezväčšia sa. Jednotlivci, ktorí dávajú mikrogaméty, sa opakovane delia a tvoria veľké číslo malé biflagelátové jedince. Mikrogaméty aktívne vyhľadávajú nehybné makrogaméty a spájajú sa s nimi a vytvárajú zygoty. Zygoty vedú k vzniku nových kolónií. Prvé dve divízie zygoty sú meiotické. V dôsledku toho má u koloniálnych bičíkovcov iba zygota diploidnú sadu chromozómov, všetky ostatné štádiá životného cyklu sú haploidné.
Objednať primárne monády (Protomonadina) Čeľaď Trypanosomy (Trypanosomatidae) Rod Leishmania (Leishmania)
Leishmania - pôvodcovia leishmaniózy - prenosné ochorenia s prírodnými ložiskami. Leishmanias objavil ruský lekár P.F.Borovský v roku 1898. Najväčší význam majú zástupcovia rodu Leishmania patriaci do čeľade Trypanozóm, ktorých rozlišovacím znakom je schopnosť vytvárať viaceré morfologicky rôzne formy v závislosti od podmienok existencie. Posun tvaru sa vyskytuje u hostiteľov bezstavovcov aj stavovcov.


Sú nasledujúce morfologické formy:
- trypanozomálna forma - charakterizovaná splošteným stuhovitým telom, v strede ktorého je oválne jadro. Bičík začína za jadrom. Axiálne vlákno bičíka smeruje k prednému koncu tela a tvorí dobre vyvinutú zvlnenú membránu. Na prednom konci tela končí a bičík vyčnieva dopredu a tvorí dlhý voľný koniec.
- Kritická forma - bičík začína mierne vpredu od jadra, pohybuje sa dopredu, tvorí krátku zvlnenú membránu a voľný koniec.
- forma leptomonas - bičík začína na samom okraji predného konca tela, nie je tam zvlnená blana, voľný koniec bičíka je značnej dĺžky.
- leishmanial forma - má zaoblený tvar a veľké okrúhle jadro. Tyčinkový kinetoplast sa nachádza na prednom konci tela. Bičík buď chýba, alebo je tam len jeho vnútrobunková časť, nepresahuje telo.
- metacyklická forma - podobná kritickej, ale bez voľného bičíka. (obr. 1).
Bičíkovce rodu Leishmania majú dve morfologické formy – leptomonas a leishmanial (obr. 2) alebo intracelulárne.
Leishmania sa delia na dermatotropné (nachádzajú sa v koži) a viscerotropné druhy (nachádzajú sa vo vnútorných orgánoch).
Pôvodca viscerálnej leishmaniózy (Leischmania donovani, L. infantum)
Lokalizácia. Bunky pečene, sleziny, kostnej drene, lymfatických uzlín, retikuloendotelové bunky podkožného tkaniva.
. Viscerálna leishmanióza je najbežnejšia v Indii (patogén L. donovani), vyskytuje sa v stredomorských krajinách, v Zakaukazsku a Strednej Ázii (patogén L. infantum), kde sa nazýva kala-azar. Vo viacerých krajinách Ázie, Afriky a Južná Amerika Ochorenia leishmaniózy sú spôsobené aj inými typmi leishmánie.
. Leptomonádové a Leishmanialove formy.

IN V poslednej dobe Zistilo sa tiež, že leishmania sídli v retikuloendoteliálnych bunkách kože, čo vysvetľuje spôsob infekcie komárov. Postihnuté bunky niekedy tvoria súvislú vrstvu alebo sú sústredené v blízkosti potných žliaz a ciev.
Patogénne pôsobenie. Existuje nepravidelná, pretrvávajúca horúčka. Slezina a pečeň sa postupne zväčšujú a môžu dosiahnuť obrovské veľkosti (obr. 3). Rozvíja sa vyčerpanie, znižuje sa obsah erytrocytov v krvi, vzniká anémia. Ochorenie môže byť akútne alebo môže mať chronický priebeh (1-3 roky). Úmrtnosť je veľmi vysoká. Väčšinou sú choré deti.
Prevencia: osobná - individuálna ochrana pred uštipnutím komárom; verejnosť - súbor opatrení na boj proti komárom a hlodavcom, ničenie túlavých a leishmanióznych psov, šakalov. Zároveň je potrebné vykonávať sanitárno-výchovné práce a ošetrovanie pacientov.
Kožná leishmanióza (Leishmania tropica)
Sú známe tri poddruhy dermatotropných druhov Leishmania: L. tropica minor a L. tropica major (na východnej pologuli) a L. tropica mexicana (na západnej pologuli).
Geografické rozloženie. Rozšírené v mnohých krajinách Európy, Ázie, Ameriky, Afriky. V Rusku sa podarilo eradikovať kožnú leishmaniózu spôsobenú L. tropica (Borovského choroba), zatiaľ čo L. major sa vyskytuje v Zakaukazsku a Strednej Ázii, najmä v mnohých regiónoch Turkménska.
Morfofyziologické charakteristiky. Leptomonas a leishmaniálne formy sú na nerozoznanie od foriem viscerotropnej leishmánie.
Životný cyklus. Takmer sa nelíši od pôvodcu viscerálnej leishmaniózy. Zdrojom nákazy sú ľudia a voľne žijúce zvieratá (drobné hlodavce žijúce v zónach piesočnatých polopúští a púští – pieskomily, sysle, škrečky, niektoré druhy potkanov a myší). Infekcia rezervoárových živočíchov v prírodných podmienkach niekedy dosahuje 70 %. Ochorenie sa u zvierat prejavuje aj vo forme kožných vredov. Komáre slúžia ako prenášače. Medzi rezervoárovými hlodavcami a vektorovými komármi je úzky kontakt. Nora hlodavcov je trvalým biotopom a živnou pôdou pre komáre, ktorých napadnutie môže dosiahnuť 35 %. Úloha človeka pri šírení kožnej leishmaniózy je malá, s výnimkou niektorých oblastí zemegule (India).
Patogénne pôsobenie. Pri kožnej leishmanióze sa tvoria zaoblené, dlhé (asi rok) nehojace sa ulcerácie na otvorené časti telo, najmä na tvári. Po zahojení zostáva znetvorujúca jazva (obr. 4). Po chorobe imunita trvá celý život.
Laboratórna diagnostika. Mikroskopické vyšetrenie výtokových vredov.
Prevencia: osobná - individuálna ochrana pred uštipnutím komárom; odporúča sa vykonať profylaktické očkovanie kmeňov kožnej leishmaniózy zo zvieraťa v uzavretých oblastiach kože. Takéto očkovanie ďalej chráni pred objavením sa znetvorujúcich vredov a jaziev na tvári a iných exponovaných častiach tela. Ako verejná prevencia sa bojuje proti komárom a hlodavcom. Najmä zavádzajú chlórpikrín do nôr, orajú pozemky obývané pieskomilmi na územiach susediacich s dedinami.

Rod Trypanosoma
Trypanosoma gambiense je pôvodcom trypanozomiázy, africkej spavej choroby u ľudí.
Lokalizácia. V tele človeka a iných stavovcov žije v krvnej plazme, lymfe, lymfatických uzlinách, cerebrospinálnom moku, tkanivách miechy a mozgu.
Geografické rozloženie. Nájdené v rovníkovej Afrike. V ohniskách ochorenia sa v krvi antilop našiel trypanozóm, ktorý slúži ako jeho prirodzený rezervoár. Nosičom je africký krv cicajúci hmyz – mucha tse-tse (Glossina palpalis). Choroba sa vyskytuje iba v distribučnej oblasti tejto muchy.
Morfofyziologické vlastnosti. Veľkosť od 13 do 39 mikrónov. Telo je zakrivené, v jednej rovine sploštené, na oboch koncoch zúžené, vybavené jedným bičíkom a zvlnenou blanou. Vyživuje sa osmoticky. Rozmnožovanie prebieha nepohlavne, pozdĺžnym delením.
Životný cyklus. Trypanosomiáza spôsobená T. gambiense je typické prenosné ochorenie s prirodzenými ložiskami. Pôvodca trypanozomiázy sa vyvíja so zmenou hostiteľa. Prvá časť životného cyklu trypanozómov prebieha v tráviacom trakte muchy tse-tse (Glossina palpalis), mimo oblasti ktorej sa trypanozomóza (africká spavá choroba) nevyskytuje, druhá časť - v tele stavovcov (hovädzí dobytok a malý dobytok, ošípané, psy, niektoré voľne žijúce zvieratá), ako aj ľudia. Keď mucha nasáva krv chorého človeka, trypanozómy sa dostanú do jej žalúdka. Tu sa množia a podstupujú sériu etáp. Celý vývojový cyklus trvá asi 20 dní. Muchy, v tele ktorých sú trypanozómy invazívneho štádia, môžu človeka nakaziť pri uhryznutí.
Patogénne pôsobenie. Pacient s trypanozomiázou má závažné lézie centrálnej nervový systém: pozorovaná svalová slabosť, vyčerpanie, duševná depresia, ospalosť. Chorobný stav trvá 7-10 rokov a ak sa nelieči, končí smrťou.
Laboratórna diagnostika. Na laboratórnu diagnostiku sa vyšetruje krv, bodky lymfatických uzlín a cerebrospinálny mok.
Prevencia. Osobná prevencia spočíva v užívaní liekov, ktoré môžu chrániť pred infekciou pri uštipnutí muchou tse-tse. Verejná prevencia spočíva v zničení nosiča. Na tento účel sa miesta rozmnožovania múch - kríkov v blízkosti obydlia a pozdĺž brehov vodných útvarov - ošetria insekticídmi alebo vysekajú.
Objednajte si polyflageláty (Polymastigina)

Črevná trichomonas (Trichomonas hominis). Spôsobuje črevnú trichomoniázu.
Lokalizácia. Dvojbodka.
Geografické rozloženie. Všade.
Morfofyziologické charakteristiky. Telo má oválny tvar so špicatým výrastkom na zadnom konci, s jedným bublinovitým jadrom. Dĺžka tela je 5-15 mikrónov, 4 voľné bičíky vybiehajú z predného konca smerom dopredu a jeden smeruje dozadu, ktorý je spojený so zvlnenou membránou. Telo je v strede prepichnuté nosnou tyčou zakončenou hrotitým hrotom na zadnom konci tela. Ústa bunky sa nachádzajú v blízkosti jadra. V cytoplazme sú tráviace vakuoly, ktoré slúžia na trávenie baktérií a črevného obsahu prehltnutého bunkou. Je možná aj osmotická výživa - tekuté látky. Rozmnožovanie je nepohlavné, pozdĺžnym delením. Schopnosť vytvárať cysty je sporná.
Infekcia sa vyskytuje cez zeleninu a ovocie, špinavé ruky kontaminované výkalmi obsahujúcimi Trichomonas, neprevarenú vodu.
Patogénne pôsobenie nie je dokázané, keďže črevná Trichomonas sa vyskytuje v črevách zdravých ľudí a pacientov s črevnými ochoreniami. Existuje názor, že črevné trichomóny nespôsobujú ochorenie, ale iba sprevádzajú patologické procesy spôsobené inými príčinami.
Laboratórna diagnostika. Mikroskopické vyšetrenie náterov výkalov, zistenie vegetatívnych foriem vo výkaloch.
Prevencia. Osobné - umývanie rúk pred jedlom a po návšteve toalety, tepelná úprava jedla a pitnej vody, dôkladné umývanie zeleniny a ovocia konzumovaného v surovom stave, ochrana potravín a vody pred prachom a muchami, ktoré môžu byť mechanickými prenášačmi rôznych patogénov. Verejná prevencia spočíva v monitorovaní hygienického stavu zdrojov vody, potravinárskych podnikov a obchodov s potravinami, verejných priestranstiev, v boji proti muchám a v šírení hygienických poznatkov.

Urogenitálny Trichomonas (Trichomonas vaginalis)- pôvodca urogenitálnej trichomonádózy.
Lokalizácia. Močové cesty mužov a žien.
Geografické rozloženie. Všade.
Morfofyziologické charakteristiky. Štruktúra je veľmi podobná črevným Trichomonas. Charakteristickými znakmi sú veľké veľkosti tela (dĺžka sa pohybuje od 7 do 30 mikrónov). Tvar tela je najčastejšie hruškovitý, sú tu 4 bičíky, zvlnená blana a nosná tyč zakončená hrotom.
Patogénne pôsobenie. Spôsobuje zápalové procesy v pohlavnom trakte, ktoré sú zdĺhavé. Predpokladá sa, že predisponujúcim momentom pre prejav patogenity je prítomnosť určitého typu baktérií v genitálnom trakte.
U žien je spočiatku postihnutá vagína, no následne sa ochorenie stáva multifokálnym. V akútnom priebehu je charakteristický hojný tekutý výtok, svrbenie a pálenie. U mužov je väčšina ochorenia asymptomatická.
Infekcia sa vyskytuje sexuálne, ako aj pri použití posteľnej bielizne, bielizne, špongie pacienta. Infekcia je možná pri vyšetrení gynekológom cez kontaminované nástroje a rukavice.
Laboratórna diagnostika. Mikroskopické vyšetrenie náterov z výtoku močových ciest. Diagnóza sa stanovuje na základe nálezu vegetatívnych foriem v sekrétoch pacienta.
Prevencia. určuje spôsob infekcie.

Lamblia (Lamblia inneris). Spôsobuje giardiázu. Tento bičíkovec objavil profesor Charkovská univerzita D. F. Lamblem v roku 1859
Lokalizácia. Dvanástnik môže po druhýkrát preniknúť do žlčovodov.
Geografické rozloženie. Všade.
Laboratórna diagnostika. Detekcia cýst vo výkaloch alebo vegetatívnych foriem v obsahu dvanástnika počas sondovania.
Prevencia. a) tepelná úprava potravín a pitnej vody; varenie zabíja cysty Giardia; b) dôkladné umytie ovocia, zeleniny a iných potravín konzumovaných v surovom stave; c) skladovanie uvarených potravín a pitnej vody v uzavretej nádobe. Treba mať na pamäti, že cysty Giardia sa môžu dostať na jedlo s prachom a zaviesť ho muchy; d) boj proti muchám. Muchy navštevujú skládky odpadových vôd, žumpy, sedia na ľudských výkaloch, hnijúcom jedle a po zafarbení tela vo výkaloch vletia do obydlia človeka, kde sedia na jedle, riade, ľudskej koži; e) umývanie rúk pred jedlom a po toalete.




