белоктың біріншілік құрылымына тән. Белоктың бастапқы құрылымы. Қосымша белок құрылымдары
Дәріс 3 Белоктардың құрылысы
Анықтамасы:
Ақуыздар - мономерлері болып табылатын тұрақты емес полимерлер Л- амин қышқылдары.
Амин қышқылдары
Табиғатта стереоизомерлердің екі түрі бар:Л (сол қолмен) және D (оң қолмен). БасқаЛ -белоктардың құрамына кіретін аминқышқылдары, организмде бар және D белоктардың құрамына кірмейтін аминқышқылдары.
Амин қышқылының жалпы формуласы суретте көрсетілген.
Бұл ақуыздарда кездесетін 20 аминқышқылдарының 19-ына қатысты. Осы 19 аминқышқылдарынан басқа белоктарда біреуі бар имино қышқылы - пролин.
Барлық аминқышқылдары бар α - амин тобы. Осыдан атауы – «α-амин қышқылдары». Пролинде - α- иминотоп.
Радикалдың полярлық (полярлық емес) принципі бойынша белоктарды құрайтын аминқышқылдарының жіктелуі.
1. Полярлы емес немесе гидрофобты радикалдар.
Алифатикалық - аланин, валин, лейцин, изолейцин.Күкірт метионин.Хош иісті - фенилаланин, триптофан.имино қышқылы пролин.
2. Полярлы, бірақ зарядсыз радикалдар. Глицин.
Оксиамин қышқылдары - серин, треонин, тирозин.құрамында сульфгидрил тобы бар цистеин.Құрамында амид тобы бар: аспарагин, глутамин.
3. Теріс зарядталған радикалдар. Аспаратин қышқылы, глутамин қышқылы.
4. оң зарядталған радикалдар. Лизин, аргинин, гистидин.
Белоктың біріншілік құрылымы
Анықтамасы:
Ақуыздың бастапқы құрылымы – полипептидтік тізбектегі аминқышқылдарының қалдықтарының реттілігі.
Аминқышқылдары коваленттік (амидтік) байланыс арқылы полипептидке қосылады.
Үш түрлі аминқышқылдарынан тұратын трипептидте 3 болуы мүмкін! = 6 түрлі бастапқы құрылымдар.
Жиырма түрлі аминқышқылдарынан тұратын олигопептидтің әртүрлі бастапқы құрылымдары 20!, бұл 2x10 18 дегенді білдіреді.
Орташа мөлшердегі ақуыздың бастапқы құрылымдарының әртүрлілігі (шамамен 500 аминқышқылдары) қазірдің өзінде шамамен 20 500 нұсқаны құрайды (егер барлық аминқышқылдары эквимолярлық қатынаста берілген болса).
ҚосулыЖер мүлдем бірдей белоктар жиынтығы бар екі адам болған емес, болмайды және болмайды.
Белоктың екіншілік құрылымы
Анықтамасы:
Ақуыздың екінші реттік құрылымы – C=O және арасындағы сутектік байланыстар есебінен полипептидтік тізбектердің реттелген құрылымы. Н- Хәртүрлі аминқышқылдары.
Екінші реттік құрылым дұрыс (α-спираль) және дұрыс емес (β-бүрмеленген құрылым) болуы мүмкін. α-спиралда NH тобы n ші амин қышқылы қалдығы (n-4)-ші амин қышқылы қалдығының C=O тобымен әрекеттеседі. Диаметрі 10,1 Å β-спиральдың бір айналымына 3,6 аминқышқылының қалдығы болады. Кәдімгі α-спиральдың идентификациялық периоды 18 амин қышқылын құрайды (5 айналым). Кәдімгі α-спиральді бұзушы бірінші кезекте пролин. Екінші маңызды әсер бірдей зарядталған, көршілес радикалдармен әсер етеді.
β-қатпарлар жалғыз ғана емес, сонымен қатар бір ақуызға кіретін іргелес полипептидтерді де құра алады.
Таза табиғи альфа немесе бета - белоктар жоқ.
Ақуыздың үшінші реттік құрылымы
Анықтама
Ақуыздың үшінші реттік құрылымы – екінші реттік құрылымы бар және радикалдар арасындағы өзара әрекеттесу арқылы анықталатын полипептидтің кеңістіктік конформациясы.
Радикалдар арасындағы өзара әрекеттесулердің төрт түрі бар.
Радикалдар арасындағы әрекеттесу түрлері
1 . ковалентті арасындағы байланыс қалдықтары екі цистеиндер (дисульфид көпірлер).
2. Қарама-қарсы зарядталған аминқышқылдарының қалдықтары арасындағы иондық (электростатикалық) әрекеттесу. («+» белгісі бар үш радикал және «-» таңбасы бар екі).
Мысалы, лизиннің оң зарядталған ε-амин тобы (- NH3 +) глутамин немесе аспарагин қышқылының теріс зарядты карбоксил тобымен - (COO-) тартылады.
3. Сутектік байланыстар.
Гидроксил, амид немесе карбоксил топтары бар барлық аминқышқылдары қатысады.
4. Гидрофобты әрекеттесулер . Полярлы емес радикалдар арасында түзілген су ортасы. 8 аминқышқылдары қатысады (бірінші класс).
Ақуыздың үшінші реттік құрылымы оның бастапқы құрылымымен толығымен анықталады, т.б. аминқышқылдарының реттілігі, ол өз кезегінде генетикалық кодпен анықталады.
Селективті емес (спецификалық емес) және көптігімен гидрофобты әрекеттесу шешуші болып табылады.
Көптеген ақуыздарда гидрофобты өзек бар.
Төрттік белок құрылымы
Анықтама: Ақуыздың төрттік құрылымы екі немесе біріктіру болып табылады Көбірекүшіншілік құрылымы бар полипептидтік тізбектер олигомерлі функционалдық маңызды құрамға айналады.
Төрттік құрылымды құрайтын және ұстап тұратын байланыстар, гидрофобтылардан басқа, үшінші реттік құрылымның қалыптасуындағымен бірдей.
Н бета тізбегінің соңы полярлы глутамин қышқылы («-» зарядталған). Орақ жасушалы анемиямен ауыратын науқастарда оның орнына полярлы емес валин қолданылады.574 аминқышқылдарының 2-і ауыстырылған.
Мұндай гемоглобин ерігіштігін жоғалтады, эритроцитті деформациялайтын талшықты тұнба түзіледі.
Орақ жасушалы анемия – генетикалық ауру. Оның себебі – гемоглобиннің В тізбегін кодтайтын гендегі бір ғана нуклеотидті ауыстыру. Бұл аллель үшін рецессивті гомозигота болып табылатын балалар екі жылға дейін өмір сүрмейді. Гетерозиготаларда 85% қалыпты және 15% ақаулы эритроциттер болады. Доминант гомозиготаларда безгек ауырады, гетерозиготаларда болмайды.
Глобулярлы және фибриллярлы белоктар
Белоктардың 95% гидрофобты ядроға ие. 5% фибриллярлық белоктар.
Глобулярлы белоктардың басым көпшілігі ериді. Фибриллярлардың көпшілігі ерімейді (α-кератиндер – шаштың, жүннің, мүйіздің, тұяқтың, тырнақтың, қабыршақтың, қауырсынның дерлік құрғақ салмағын құрайды; коллаген – сіңір ақуызы, шеміршек; фиброин – жібек ақуызы).
Фибриллярлы белоктарда глобулярлыларға қарағанда зарядталған аминқышқылдарының көп үлесі бар - жеке тізбектер ериді, ал олардың кешендері полюссіз және ерімейді.
ПРОТЕИНДІҢ ҚҰРЫЛЫМЫ
Белоктардың құрылымында молекуланың ұйымдастырылуының төрт деңгейі бөлінеді: біріншілік, екіншілік, үшіншілік және төрттік құрылымдар. Алғашқы екі деңгей барлық белоктарға тән. Үшіншілік және төрттік құрылымдар тек глобулярлы белоктарда болады.
Белоктардың біріншілік құрылымы
|
Пептидтік байланыстың түзілуі |
Белоктардың бастапқы құрылымы – полипептидтік тізбектегі аминқышқылдарының қалдықтарының реттілігі. Ақуыздағы аминқышқылдарының орналасу реті ДНҚ-дағы нуклеотидтердің реттілігімен генетикалық түрде анықталады. Полипептид бір амин қышқылының карбоксил тобының екінші амин қышқылының амин тобымен – пептидтік байланыспен әрекеттесуінен түзіледі.
Бір амин қышқылының «басы» (NH2-) екінші амин қышқылының «құйрығына» (-COOH) бекітіледі. Аминқышқылдары арасында пептидтік байланыс (-CO-NH-) тұйықталған, бұл ақуыздың біріншілік құрылымындағы байланыстың жалғыз түрі. Жоғарыдағы сұлбадан көрініп тұрғандай, пептидтік байланыстың түзілуі кезінде су бөлінеді. Гидролиз кезінде пептидтік байланыстың ыдырауы ыдырайтын байланыс орнында судың қосылуымен бірге жүреді. Ақуыздар мен полипептидтердің гидролизінің соңғы өнімі бос аминқышқылдары болып табылады.
Пептидтік байланыс көміртек пен азот арасындағы жалғыз байланысқа қарағанда күшті, өйткені таутомерия нәтижесінде ол 40% қосарланады. Сол себепті полипептидтік тізбекте айналу тек радикалмен байланысқан көміртегі атомдарының айналасында ғана мүмкін.
Барлық полипептидтердің қаңқасы бірдей. Полипептидтік тізбектер табиғаты мен радикалдарының реттілігі бойынша ерекшеленеді. Полипептидті оның құрамына кіретін аминқышқылдарының қалдықтарының санына қарай атайды: дипептид, трипептид және т.б.
Белоктар – құрамында 50-ден астам амин қышқылы қалдықтары бар полипептидтер. Ең қарапайым ақуыз - инсулин. Оның құрамында тек 51 амин қышқылының қалдығы бар. Рибонуклеазаға 124 қалдық, гемоглобин 574 кіреді.
Белоктарда амин қышқылдарының реті, яғни біріншілік құрылымы қатаң анықталған. Бір аминқышқылының қалдықтарын екіншісімен алмастыру жаңа ақуызды шығарады. Сонымен, сиыр инсулинінің тоғызыншы позициясында серин қалдығы, ал қошқар инсулинінде - глицин бар. Адам мен жылқы инсулиніндегі айырмашылықтар үш аминқышқылының қалдықтарына қатысты - сегізінші, тоғызыншы және оныншы. Барлық аталған инсулиндердің бастапқы құрылымы әртүрлі. Атқаратын қызметі бірдей әр түрлі организмдердің белоктары гомологиялық деп аталады.
Белоктардың екіншілік құрылымы
Ақуыздарда екінші реттік құрылымның екі негізгі түрі бар: спираль және бүктелген қабат.
Спиральдар . Полипептидтік тізбектегі α-көміртек атомының айналасындағы байланыстардың еркін айналуына байланысты полипептидтік тізбектің сызықтылығы бұзылады. Бұл спиральдардың пайда болуына әкеледі. Спиральдың 3 түрі бар.
1. Кератинмен сипатталады α-c спираль. Кератиннің полипептидтік тізбегі ойдан шығарылған цилиндрге оралған сияқты. Бір-біріне жақын орналасқан катушкалар сутегі байланыстарымен ұсталадыбір пептидтік байланыстың оттегі мен екінші пептидтік байланыстың сутегі арасында. Сутектік байланыстар оттегі мен сутегі арасындағы коваленттік байланыстардан 20 есе әлсіз, бірақ олардың көптігіне байланысты олар спиральді өте берік ұстайды.
2. β -спиральбактериялық ақуыздарда кездеседі. p-спиралының бір айналымы 22 аминқышқылының қалдықтарынан тұрады, β-спираль қуыс түтік, ал α-спираль толтырылған цилиндр.
3. сынған спиральколлагенге тән. Спиральдың бұл түрі коллагендегі гидроксипролині бар глицин мен пролиннің жоғары құрамының салдары - спиральдың «дұрыстығын» бұзатын аминқышқылдары.
МЕН бүктелген қабат жібек протеиніне тән – фиброин. Бүктелген қабаттағы іргелес тізбектердің бағыты қарама-қарсы (антипараллель) Бір-біріне жақын тізбектер сутегі байланыстарымен ұсталады.
Фибриллярлық протеиндердегі спиральдар мен қатпарлы қабаттар көбінесе суперекінші құрылымдарды немесе супер орамдарды тудырады. Сонымен, кератиннің 7 α-спиральдары супер орамды құрайды. Өз кезегінде 11 кератинді суперкоил шаш микрофибрилін құрайды.
Глобулярлы белоктардың екіншілік құрылымы фибриллярлық белоктар сияқты біркелкі емес. Сонымен, миоглобин молекуласында полипептидтік тізбектің 77%-ы спиральданған, ал 23%-ы спиральданбаған. Инсулиннің спиральдану дәрежесі - 60%, жұмыртқа альбумині - 40%, пепсин - 28%. Химотрипсиннің полипептидтік тізбегінде спиральданған бөліктер жоқ дерлік, бірақ қатпарлар, қабаттар, ілмектер, иілулер және т.б.
Молекулярлық салмағы 20 мыңнан асатын глобулярлы белоктардың құрылымында Иә, концепция ерекшеленеді. домен - өзіне тән құрылымы бар 100-150 аминқышқылдарының қалдықтарының шағын аймақтары. Олар құрылымдық домендер деп аталады.
Домендер мен жеке тұлғалар арасында құрылыс блоктарыдомен ішінде деп аталатындар бар топсалы бөлімдер . Көбінесе бір ақуызда бір типті бірнеше ұқсас домендер кездеседі.
Басқа тұжырымдама бар функционалдық домен . Соңғы жағдайда бір немесе бірнеше құрылымдық домендер бірігіп белок молекуласында функционалды түрде бөлек учаскені құрайды: субстрат алаңы, ферменттің немесе ингибитордың белсенді орталығының ортасы, мембранадағы иондық канал және т.б.
Үшіншілік құрылым- үш өлшемді кеңістікте полипептидтік тізбектің орналасуы (спирализацияланған, төмен орамды немесе орамсыз).
Глобулярлы катушканың көрінетін бұзылуына қарамастан, оның құрылымы қатаң анықталған және кейбір заңдылықтарға ие.
1. Глобулдағы полипептидтік тізбектер өте тығыз оралған.
2. Әдетте белоктың полярлық топтары глобуланың бетінде орналасады, ал оның ішінде гидрофобты радикалдар жасырылады.
Ацетон" href="/text/category/atceton/" rel="bookmark">Ацетон ақуызы тұнбаға түседі. Бұл тұнба деп аталады. тұздану. Тұздану механизмі - тұз иондары мен спирт пен ацетон молекулалары, күшті гидратация қабығы бар, суды ақуыз молекуласынан алады. Тұздардың әртүрлі концентрацияларында әртүрлі ақуыздар тұзданады. Глобулиндер аммоний сульфатының жартылай қаныққан ерітіндісінде, ал альбуминдер тек осы тұздың қаныққан ерітіндісінде тұзданады. Фракциялық тұздану ақуыздарды бөлу және тазарту үшін қолданылады.
Кейбір ақуыздар сәйкес рН-да тұнбаға түседі изоэлектрлік нүкте. Осылайша, казеин рН 4,7-де тұнбаға түседі, өйткені бұл рН кезінде молекулалардың заряды болмайды және ерітіндіде тұрақсыз ірі бөлшектерге тез біріктіріледі. Басқа белоктар тұрақтырақ, олардың тұнбаға түсуі үшін ақуыз тұрақтылығының екі факторына да әсер ету қажет.
Протеин диализі
Үлкен өлшемдерге байланысты ақуыз молекулалары кейбір пленкалар арқылы енбейді; целлофан, балық көпіршіктері және т.б. Бұл қасиет белоктарды төмен молекулалық қоспалардан тазарту үшін, яғни диализ үшін пайдаланылады.
Тұздар қоспасы бар ақуыз ерітіндісі полиэтилен пакетіне құйылады, сөмке тазартылған су ағатын ыдысқа салынады. Тұздардың және басқа заттардың ұсақ иондары целлофанға суға еніп, жойылады, ал ақуыз ерітіндісі қапта қалады.
ақуыз заряды
Белоктың құрамында, әдетте, қышқыл, теріс зарядталған амин қышқылдарының қосындысы (глютамин, аспарагин) негізгі, оң зарядталған амин қышқылдарының (лизин, аргинин, гистидин) қосындысына тең емес. Осыған байланысты судағы ақуыздардың заряды оң немесе теріс болады. Мұндай ақуыздың ерітіндісін қышқылдандырғанда (Н+ артық) карбоксил топтарының иондануы басылып, оң зарядты топтардың қосындысы теріс зарядтылардың қосындысына тең болатын сәт келеді. Бұл жағдайда ақуыз молекуласы тұтастай зарядсыз болады. Ақуыздың бұл күйі деп аталады ойэлектрлік, ал изоэлектрлік күй пайда болатын рН изоэлектрлік нүкте (ИЭП) деп аталады. IEP - ақуыздың ең маңызды сипаттамаларының бірі.
Ерітіндіні одан әрі қышқылдандыру кезінде ақуыз оң зарядталады. Ақуыз молекулаларының қайта зарядталуы жүреді. Оң зарядталған ақуызды алсақ, онда сілтіленгенде ол алдымен изоэлектрлік күйге ие болады, содан кейін теріс зарядталады.
Жалпы ереже: IEP төмен рН-дағы ақуыз оң зарядталған және катион, ал IEP-тен жоғары рН кезінде ол теріс зарядталған және анион болып табылады.
Белоктардың зарядтарының айырмашылығы оларды тұрақты электр өрісінде бөлуге мүмкіндік береді. Бұл бөлу әдісі электрофорез деп аталады.
Ион алмасу хроматографиясы сонымен қатар қоспаның бөлінген заттардың зарядтарының айырмашылығына негізделген.
Ақуыздың денатурациясы
Денатурация - белоктардың құрылымындағы гидролитикалық емес өзгерістер, олардың өзгеруімен бірге жүреді. биологиялық белсенділікжәне функциялары. Көптеген факторлар денатурацияны тудыруы мүмкін: қайнау, жоғары температура, ультракүлгін және иондаушы сәулелену, артық қысым, ауыр металдардың тұздары, экстремалды рН мәндері (күшті қышқылдар мен сілтілер), кейбір органикалық қосылыстар.
Қыздыру және әртүрлі сәулелену белоктағы сутегі мен иондық байланыстарды бұзады. Күшті қышқылдар, сілтілер және концентрлі тұз ерітінділері иондық байланыстарды бұзады. Ауыр металдар карбоксианиондармен күшті байланыс түзеді және иондық байланыстарды үзеді. Органикалық еріткіштер мен жуғыш заттар гидрофобты әрекеттесулерді бұзады және ақуыздардағы сутектік байланыстарды бұзады.
Денатурация кезінде белоктың барлық әлсіз байланыстары өзгереді немесе жойылады: сутегі, электростатикалық, гидрофобты және т.б., бірақ пептидтік байланыстар өзгеріссіз қалады.
Денатурацияның белгілері:
1) ерігіштіктің өзгеруі. Суда еріген ақуыз тұнбаға түседі немесе керісінше, ерімейтін ақуыз ерітіндіге түседі;
2) оптикалық белсенділіктің өзгеруі, мысалы, поляризацияланған сәуленің жазықтығының айналу бұрышы;
3) белок глобулының ішінде денатурация алдында жасырылған жаңа реактивті топтардың пайда болуы;
4) денатурацияның негізгі және бірінші белгісі – функцияның жоғалуы. Құрылымдық ақуызборпылдақ болады, ферменттер каталитикалық белсенділігін жоғалтады және т.б.
Денатурациялаушы агенттен бөлінгеннен кейін белок біртіндеп өзінің бастапқы қасиеттеріне ие болады. Бұл процесс деп аталады ренатурация.
Белоктардың оптикалық қасиеттері
Хромопротеидтерден басқа белоктар түссіз. Белоктар ультракүлгін сәулелерді хош иісті аминқышқылдарына байланысты λ=280 нм максимуммен жұтады. λ=216 нм кезіндегі екінші абсорбция максимумы пептидтік байланысқа жатады.
Протеин ерітінділері мөлдір, бірақ опалесценцияға ие - бүйірлік жарықтандыру астында бұлыңғырлық көрінеді. Бұл қасиеттер ақуыздың мөлшерін анықтау үшін қолданылады.
МОНОНУКЛЕОТИДТЕР


Пурин Гуанин Аденин
Пиримидин цитозин тимин урацил
Көрсетілген негіздермен қатар, азотты негіздердің метилденген, құрамында күкірт бар және басқа да туындылары бар. Олар деп аталады кіші негіздер. Мысалы, прокариоттарда: риботимидин, инозин, ксантин, гипоксантин және т.б. Барлығы 60-қа жуық азотты негіздер белгілі.
Азотты негіздер мен олардан құрастырылған қосылыстар ультракүлгін аймақта (260-280 нм) жарықты қарқынды сіңіреді. Бұл қасиет олардың құрамында азотты негіздер бар заттарды сандық анықтау үшін қолданылады.
https://pandia.ru/text/78/240/images/image009_58.jpg" alt="http://*****/biohimija_severina/img/B5873p267-a1.jpg" align="left" width="289" height="203 src=">Важным производным нуклеозидов является !} cAMP. Ол аденилатциклаза ферментінің қатысуымен АТФ-дан түзіледі. cAMP жасушадағы зат алмасу процестерін реттеуге қатысады. Атап айтқанда, ол белгілі бір гормондардың жасушаға әсер етуінде екінші медиатор қызметін атқарады. .
Нуклеотидтердің түріне сәйкес құрастырылған қосылыстар кейбір күрделі ферменттердің құрамына кіреді, рөл атқарады коферменттер. Көбінесе мұндай коферменттердің құрамында пуриндік және пиримидиндік негіздерден құрылымы жағынан ерекшеленетін азотты заттар кездеседі. Олар жануарлардың ағзасында синтезделмейді, бірақ тамақтан (витаминдерден) пайда болады.
Флавин мононуклеотиді ( FMN) - фосфорланған рибофлавин(В2 дәрумені).
Флавин аденин динуклеотиді ( FAD) екі нуклеотидтен AMP және FMN тұрады.

Коэнзим А тиоэфир байланысы арқылы SH тобына қосылған ацил радикалдарын белсендіреді және тасымалдайды.
Қышқылға төзімділікке байланысты қосылыстар: ацетил-кофермент А, малонил-кофермент А, сукцинил-коэнзим А деп аталады.
НУКЛЕИН Қышқылдары
Нуклеин қышқылдары- дезоксирибонуклеиндік (ДНҚ) және рибонуклеиндік (РНҚ) сәйкесінше дезоксирибонуклеотидтер мен рибонуклеотидтердің полимерлері. Нуклеин қышқылдарының мононуклеотидтері фосфор қышқылының қалдығы арқылы байланысады. рибозаның бесінші көміртегіЖәне рибозаның үшінші көміртегікөрші нуклеотид.
ДНҚ құрылымы
1950 жылы Чаргафф ДНҚ-ның нуклеотидтік құрамының бірқатар заңдылықтарын ашты, кейін олар Чаргафф ережелері деп аталды. Бұл ережелер: 1) Pur=Pir, 2) A=T, 3) G=C, 4) A+C=G+T. Чаргафф ережелері ДНҚ құрылымының қосымша моделін құруға көмектесті.
ДНҚ-ның бастапқы құрылымыполинуклеотидтік тізбектермен бейнеленген.
|
 ДНҚ құрылымы
ДНҚ құрылымыДНҚ тізбектерінде бірнеше үлгілер бар:
1) Вирустар мен прокариоттарда ДНҚ тізбегі түгел дерлік бірегей, эукариоттарда ДНҚ-ның 30-40% қайталанатын тізбектер, әсіресе центромера аймағындағы ДНҚ-ның көптеген қайталанатын бөлімдері.
2) ДНҚ тізбектерінің тармақтары болмайды.
3) ДНҚ-да көптеген (мыңдаған) кері жүретін тізбектер бар – палиндромдар, «өзгерістер». Орыс тіліндегі ауыстырғыштардың мысалдары: «баклажанға қабанды басты». Палиндромдар ген экспрессиясын (жұмысын) реттеуде маңызды рөл атқаратын крест тәрізді құрылымдарды – шаш қыстырғыштарын құрайды.
ДНҚ-ның екіншілік құрылымы
1953 жылы Дж.Уотсон мен Ф.Крик ДНҚ қос спираль екенін анықтады. антипараллельполинуклеотидтік тізбектер. Тізбектер азотты негіздер арасында түзілген сутектік байланыстар арқылы бір-біріне жақын ұсталады, ал аденин мен тимин арасында екі есе, цитозин мен гуанин арасында үш есе байланыс бар. Сыртта қос спиральДНҚ-да қант фосфатты діңгегі бар.
Қосымша байланысқан азотты негіздер ішке қарайды. Стекте азотты негіздер бір-біріне қатысты ығысады. ДНҚ спиральының бірнеше түрі бар:
1) ДНҚ репликациясы кезінде табылған В типті спираль;
2) транскрипция кезінде байқалатын А типті спираль;
3) А немесе В спиралі сияқты оңға емес, солға бұралған Z типті спираль қиылысу кезінде пайда болады.
4) C және SBS типті катушкалар да сипатталған. Соңғысы жабық емес.
Вирустарда бір тізбекті ДНҚ болуы мүмкін.
ДНҚ-ның үшінші реттік құрылымы
Прокариоттарда дөңгелек ДНҚ молекулалары болады. Эукариоттарда ДНҚ ұштары бос – бұл ДНҚ-ның сызықтық түрі. Вирустарда сызықтық және шеңберлік ДНҚ болады.
Прокариоттарда ядро болмайды. Олардың ДНҚ белоктармен бірге цитоплазмалық мембранаға бекітіліп, нуклеоид түзеді.
Эукариоттарда ДНҚ жасушаның қалған бөлігінен ядролық мембрана арқылы бөлінеді. Интерфазада эукариоттық ДНҚ шоғырланған хроматин жіптері. Хроматин құрамында ДНҚ-дан басқа белоктар бар. 50% хроматин ақуызы - гистондар. Гистондар құрамында үлкен сандиаминокарбон қышқылдарының қалдықтары: аргинин және лизин. Бұл барлық эукариоттарда бірдей дерлік өте сақталған глобулярлы белоктар. Хроматин белоктарының екінші жартысы үлкен әртүрлілікпен сипатталатын гистонды емес ақуыздар болып табылады.
 Хроматиннің бірнеше ұйымдасу деңгейі бар:
Хроматиннің бірнеше ұйымдасу деңгейі бар:
1) Нуклеосомалар. ДНҚ-ның екі дерлік айналымы төрт жұп гистон молекуласының өзегіне оралған. Бұл - негізгі. Кортекс арасында орналасқан байланыстырушы- ішінара гистон және (немесе) гистон емес ақуыздармен жабылған немесе ақуыздармен мүлде қамтылмаған 40 негіз жұбы. Гистондар транскрипциялық деңгейде гендердің активтенуіне және репрессиясына қатысады.
2) соленоидтар: 6-10 нуклеосома соленоидтың бір айналымын жасайды.
3) ілмектер. Гистонды емес белоктардың қаңқасында 30-90 мың жұп негізді ілмектер кездеседі, олардың басы мен соңы жақын орналасқан.
4) Жоғарғы деңгейЭукариоттарда ДНҚ-ның ұйымдастырылуы хромосома болып табылады. Хромосоманың негізін ДНҚ бекітілген белок матрицасы құрайды. Хромосоманың ұштарында ДНҚ бөлімдері деп аталады теломерлер. Репликация теломерлерден басталуы мүмкін; теломерлер хромосомалардың ұштарын деградациядан қорғайды.
Әрбір репликация кезінде теломерлер қысқарады. Теломералардың белгілі бір сыни шағын мәніне жеткеннен кейін жасуша өледі. Теломераза – теломерлердің ұзындығын қалпына келтіретін, жасушаны өлмейтін ететін фермент. Теломераза репродуктивті, сабақта кездеседі Жәнерак клеткалары, басқа жасушалар жоқ. Хромосоманың ортасында орналасқан центромер- сонымен қатар жасушаның бөлінуі кезінде хромосомалардың дұрыс дивергенциясын қамтамасыз ететін кодталмаған ДНҚ.
ДНҚ-ның көп бөлігі ілмектерде. Бұл жерде гендер орналасқан. Әрбір циклде бір немесе бірнеше ген бар. Циклдер хромосомалық матрицамен ДНҚ кодталмаған аймақтары арқылы әрекеттеседі.
ДНҚ-ның физикалық және химиялық қасиеттері
Хромосома - бұл бір ДНҚ молекуласы. Прокариоттарда бір ғана хромосома болады. ДНҚ өлшемдері вирустардағы 5000 нуклеотидтен адамдарда 5 миллиардқа дейін (оның ұзындығы 8 см) өзгереді. Адамның бір жасушасының барлық хромосомаларының ДНҚ ұзындығы шамамен 2 м.
ДНҚ – ақ талшықты масса. Ерітінділер өте тұтқыр. Молекулярлық массаның ұлғаюымен тұтқырлық артады. ДНҚ ерітінділері ультракүлгін сәулелерді максималды 260 нм-де сіңіреді. Судағы ДНҚ теріс зарядты.
Қышқыл, сілтілі ортада ° C температурада формамид, мочевина және басқа да бірқатар факторлардың қатысуымен ДНҚ полинуклеотидтік тізбектерінің дивергенциясы пайда болады - денатурация. Денатурация кезінде сутектік байланыстар үзіледі - ДНҚ » ериді". Балқу температурасы ДНҚ жартылай денатурацияланатын температура деп саналады (сутектік байланыстың жартысы үзіледі). Балқу кезінде ерітінділердің оптикалық тығыздығының 260 нм жоғарылауы байқалады - гиперхромды әсер.
ДНҚ-да G-C жұптары неғұрлым көп болса, балқу температурасы соғұрлым жоғары болады, өйткені G-C жұптары A-T қарағанда күштірек, өйткені олар үш сутектік байланыспен ұсталады.
Температура төмендегеннен кейін жылумен денатурацияланған ДНҚ қайталама құрылымын қалпына келтіреді, ренатурация жүреді немесе күйдіру, қышқылдар.
Егер қоспадағы әр түрлі көздерден алынған ДНҚ денатурацияға және жасытуға ұшыраса, онда бөгде ДНҚ тізбектерінің гибридтенуі комплементарлық заңдар бойынша жүреді. ДНҚ және РНҚ тізбектерінің гибридтенуі мүмкін. Бұл жағдайда гибридті нуклеин қышқылымұндағы бір тізбек РНҚ, ал екінші тізбек ДНҚ.
1-МОДУЛЬ НЕГІЗДІҢ ҚҰРЫЛЫМЫ, ҚАСИЕТТЕРІ ЖӘНЕ ҚЫЗМЕТІ
1-МОДУЛЬ НЕГІЗДІҢ ҚҰРЫЛЫМЫ, ҚАСИЕТТЕРІ ЖӘНЕ ҚЫЗМЕТІ
Модуль құрылымы | Тақырыптар |
Модульдік блок 1 | 1.1. Белоктардың құрылымдық ұйымдасуы. Белоктардың нативті конформациясының қалыптасу кезеңдері 1.2. Белоктардың қызмет ету негіздері. Белок қызметіне әсер ететін лигандтар ретіндегі препараттар 1.3. Ақуыздың денатурациясы және олардың өздігінен ренатациялану мүмкіндігі |
Модульдік блок 2 | 1.4. Гемоглобин мысалында олигомерлі белоктардың құрылысы мен қызметінің ерекшеліктері 1.5. Жасушадағы белоктардың нативті конформациясын сақтау 1.6. Белоктардың әртүрлілігі. Иммуноглобулиндер мысалында белоктар тұқымдастары 1.7. Белоктардың физика-химиялық қасиеттері және оларды бөлу әдістері |
Модульдік блок 1 МОНОМЕРЛІ БЕЛГІЛЕРДІҢ ҚҰРЫЛЫМДЫҚ ҰЙЫМДАСТЫРЫЛУЫ ЖӘНЕ ОЛАРДЫҢ ҚЫЗМЕТІНІҢ НЕГІЗДЕРІ
Оқу мақсаттары Істей білу:
1. Тұқым қуалайтын және жүре пайда болған протеинопатиялардың даму механизмдерін түсіну үшін белоктардың құрылымдық ерекшеліктері және белок функцияларының олардың құрылымына тәуелділігі туралы білімдерін пайдалану.
2. Белоктармен әрекеттесіп, белсенділігін өзгертетін лигандтар ретіндегі кейбір дәрілік заттардың емдік әсер ету механизмдерін түсіндіріңіз.
3. Белоктардың құрылымы мен конформациялық лабильділігі туралы білімдерін олардың құрылымдық және функционалдық тұрақсыздығын және өзгермелі жағдайларда денатурацияға бейімділігін түсіну үшін пайдаланыңыз.
4. Медициналық материал мен аспаптарды, сондай-ақ антисептиктерді зарарсыздандыру құралы ретінде денатурациялаушы заттарды қолдануды түсіндіріңіз.
Білу:
1. Белоктардың құрылымдық ұйымдасу деңгейлері.
2. Белоктардың құрылымдық және қызметтік әртүрлілігін анықтайтын біріншілік құрылымының маңызы.
3. Белоктардағы белсенді орталықтың түзілу механизмі және оның белоктар қызметінің негізінде жатқан лигандпен спецификалық әсерлесуі.
4. Экзогендік лигандтардың (дәрілік заттар, токсиндер, улар) белоктардың конформациясы мен функционалдық белсенділігіне әсер ету мысалдары.
5. Белок денатурациясының себептері мен салдары, денатурацияны тудыратын факторлар.
6. Медицинада денатурациялаушы факторларды антисептиктер және медициналық аспаптарды зарарсыздандыруға арналған құралдар ретінде қолдану мысалдары.
ТАҚЫРЫП 1.1. БЕЛГІЛЕРДІҢ ҚҰРЫЛЫМДЫҚ ҰЙЫМДАСТЫРЫЛУЫ. ТУҒАН ТҰРҒАНЫН ҚАЛЫПТАСТЫРУ КЕЗЕҢДЕРІ
ПРОТЕИНДІҢ КОНФОРМАЦИЯЛАРЫ
Ақуыздар - полимерлі молекулалар, олардың мономерлері тек 20 α-аминқышқылдары. Белоктағы амин қышқылдарының жиынтығы мен байланысу реті даралардың ДНҚ-дағы гендердің құрылымымен анықталады. Әрбір белок өзінің ерекше құрылымына сәйкес өз қызметін атқарады. Белгілі бір организмнің белоктарының жиынтығы оның фенотиптік ерекшеліктерін, сондай-ақ тұқым қуалайтын аурулардың болуын немесе олардың дамуына бейімділігін анықтайды.
1. Белоктарды құрайтын аминқышқылдары. пептидтік байланыс.Белоктар – мономерлерден – 20 α-аминқышқылдарынан түзілген полимерлер, олардың жалпы формуласы:
Аминқышқылдары α-көміртек атомына қосылған радикалдардың құрылымы, мөлшері, физика-химиялық қасиеттері бойынша ерекшеленеді. Аминқышқылдарының функционалдық топтары әртүрлі α-аминқышқылдарының қасиеттерінің ерекшеліктерін анықтайды. α-аминқышқылдарының құрамындағы радикалдарды бірнеше топқа бөлуге болады:
пролин,Басқа 19 ақуыз мономерлерінен айырмашылығы, амин қышқылы емес, иминоқышқыл, пролиндегі радикал α-көміртек атомымен де, имино тобымен де байланысты.
 Аминқышқылдары суда ерігіштігімен ерекшеленеді.Бұл радикалдардың сумен әрекеттесу қабілетіне (гидратациялануға) байланысты.
Аминқышқылдары суда ерігіштігімен ерекшеленеді.Бұл радикалдардың сумен әрекеттесу қабілетіне (гидратациялануға) байланысты.
TO гидрофильдіқұрамында аниондық, катиондық және полярлық зарядсыз радикалдар жатады функционалдық топтар.
TO гидрофобтықұрамында метил топтары, алифаттық тізбектер немесе циклдар бар радикалдар жатады.
2. Пептидтік байланыс аминқышқылдарын пептидтерге байланыстырады.Пептид синтезі кезінде бір амин қышқылының α-карбоксил тобы екінші амин қышқылының α-амин тобымен әрекеттеседі. пептидтік байланыс:
Белоктар полипептидтер, яғни. байланысқан α-аминқышқылдарының сызықтық полимерлері пептидтік байланыс(Cурет 1.1.)
 Күріш. 1.1. Пептидтердің құрылысын сипаттауда қолданылатын терминдер
Күріш. 1.1. Пептидтердің құрылысын сипаттауда қолданылатын терминдер
Полипептидтерді құрайтын аминқышқылдарының мономерлері деп аталады аминқышқылдарының қалдықтары.Қайталанатын топтар тізбегі - NH-CH-CO- пішіндер пептидтік магистраль.Бос α-амин тобы бар аминқышқылдарының қалдығы N-терминал деп аталады, ал бос α-карбоксил тобы бар аминқышқылдарының қалдығы С-терминал деп аталады. Пептидтер N-терминусынан С-соңына дейін жазылады және оқылады.
Пролиннің имино тобынан түзілетін пептидтік байланыс басқа пептидтік байланыстардан ерекшеленеді: пептидтік топтың азот атомында сутегі,
оның орнына радикалмен байланыс бар, нәтижесінде циклдің бір жағы пептидтік магистральға кіреді:
 Пептидтер аминқышқылдарының құрамымен, аминқышқылдарының санымен және аминқышқылдарының орналасу ретімен ерекшеленеді, мысалы, Сер-Ала-Глу-Гис және Хис-Глу-Ала-Сер екі түрлі пептидтер.
Пептидтер аминқышқылдарының құрамымен, аминқышқылдарының санымен және аминқышқылдарының орналасу ретімен ерекшеленеді, мысалы, Сер-Ала-Глу-Гис және Хис-Глу-Ала-Сер екі түрлі пептидтер.
Пептидтік байланыстар өте күшті және олардың химиялық ферментативті емес гидролизі үшін қатал жағдайлар қажет: талданатын ақуыз концентрлі тұз қышқылында шамамен 110°С температурада 24 сағат бойы гидролизденеді. Тірі жасушада пептидтік байланыстарды бұзуға болады протеолитикалық ферменттер,шақырды протеазаларнемесе пептидті гидролазалар.
3. Белоктардың біріншілік құрылымы.Әртүрлі белоктардың пептидтік тізбектеріндегі аминқышқылдарының қалдықтары ретсіз кезектеспей, белгілі бір ретпен орналасады. Полипептидтік тізбектегі аминқышқылдары қалдықтарының сызықтық тізбегі немесе тізбегі деп аталады. белоктың бастапқы құрылымы.
Әрбір жеке ақуыздың бастапқы құрылымы ДНҚ молекуласында (ген деп аталатын аймақта) кодталған және транскрипция (мРНҚ туралы ақпаратты қайта жазу) және трансляция (ақуыздың бастапқы құрылымының синтезі) кезінде жүзеге асырылады. Демек, жеке адамның белоктарының бастапқы құрылымы ата-анадан балаларға мұра болып табылатын ақпарат болып табылады, ол белгілі бір ағзаның белоктарының құрылымдық ерекшеліктерін анықтайды, оған бар белоктардың қызметі байланысты болады (1.2-сурет).
 Күріш. 1.2. Жеке адам ағзасында синтезделген белоктардың генотипі мен конформациясы арасындағы байланыс
Күріш. 1.2. Жеке адам ағзасында синтезделген белоктардың генотипі мен конформациясы арасындағы байланыс
Адам ағзасындағы шамамен 100 000 жеке белоктың әрқайсысында бар бірегейбастапқы құрылым. Белоктың бір түрінің молекулалары (мысалы, альбумин) альбуминді кез келген басқа жеке белоктан ерекшелендіретін аминқышқылдарының қалдықтарының бірдей кезектесуіне ие.
Пептидтік тізбектегі аминқышқылдары қалдықтарының реттілігін ақпаратты тіркеу формасы ретінде қарастыруға болады. Бұл ақпарат сызықтық пептидтік тізбектің кеңістіктік қатпарлануын анықтайды, үш өлшемді құрылым деп аталады. конформациятиін. Функционалды белсенді белок конформациясының түзілу процесі деп аталады бүктеу.
4. Белоктардың конформациясы.Пептидтік магистральда еркін айналу пептидтік топтың азот атомы мен көршілес α-көміртек атомы арасында, сондай-ақ α-көміртек атомы мен карбонил тобы көміртегі арасында мүмкін. Аминқышқылдары қалдықтарының функционалдық топтарының өзара әрекеттесуіне байланысты белоктардың біріншілік құрылымы күрделірек кеңістіктік құрылымдарға ие болуы мүмкін. Глобулярлы белоктарда пептидтік тізбектердің конформациясының қатпарлануының екі негізгі деңгейі бөлінеді: қосалқыЖәне үшінші құрылым.
Белоктардың екіншілік құрылымы- бұл пептидтік магистральдың -C=O және -NH- функционалдық топтары арасында сутектік байланыстардың түзілуі нәтижесінде түзілетін кеңістіктік құрылым. Бұл жағдайда пептидтік тізбек екі типті тұрақты құрылымдарды ала алады: α-спиральдарЖәне β құрылымдар.
IN α-спиральдаркарбонил тобының оттегі атомы мен одан 4-ші амин қышқылының амидті азотының сутегі арасында сутектік байланыстар түзіледі; амин қышқылы қалдықтарының бүйірлік тізбектері
екінші реттік құрылымның түзілуіне қатыспай, спиральдың шеткі жағында орналасқан (1.3-сурет).
Бірдей зарядты алып жүретін көлемді радикалдар немесе радикалдар α-спиралдың түзілуіне жол бермейді. Сақина құрылымы бар пролин қалдығы α-спиралды үзеді, өйткені пептидтік тізбектегі азот атомында сутегінің болмауына байланысты сутегі байланысын құру мүмкін емес. Азот пен α-көміртек атомы арасындағы байланыс пролиндік циклдің бөлігі болып табылады, сондықтан пептидтік магистраль осы жерде иілуге ие болады.
β-құрылымыбір полипептидтік тізбектің пептидтік магистралінің сызықтық аймақтары арасында түзіледі, осылайша қатпарлы құрылымдарды құрайды. Полипептидтік тізбектер немесе олардың бөліктері түзілуі мүмкін параллельнемесе параллельге қарсы β-құрылымдар.Бірінші жағдайда әрекеттесетін пептидтік тізбектердің N- және С-терминалдары сәйкес келеді, ал екінші жағдайда олар қарама-қарсы бағытта болады (1.4-сурет).
Күріш. 1.3. Белоктың екінші реттік құрылымы – α-спираль
 Күріш. 1.4. Параллельді және антипараллельді β бүктемелі құрылымдар
Күріш. 1.4. Параллельді және антипараллельді β бүктемелі құрылымдар
β-құрылымдар кең көрсеткілермен белгіленеді: A - антипараллель β-құрылым. B - параллель β-бүрмеленген құрылымдар
Кейбір белоктарда β-құрылымдар әртүрлі полипептидтік тізбектердің пептидтік магистраль атомдары арасында сутектік байланыстардың түзілуіне байланысты түзілуі мүмкін.
Ақуыздарда да кездеседі тұрақты емес қайталама аймақтарқұрылым, оның құрамына полипептидті омыртқаның иілісі, ілмектері, бұрылыстары кіреді. Олар көбінесе пептидтік тізбектің бағыты өзгеретін жерлерде орналасады, мысалы, параллель β-парақ құрылымын қалыптастыру кезінде.
α-спиральдар мен β-құрылымдардың болуы бойынша глобулярлы белоктарды төрт категорияға бөлуге болады.
Күріш. 1.5. Миоглобин (А) мен гемоглобин β-тізбегінің (В) екінші реттік құрылымы, құрамында сегіз α-спираль бар

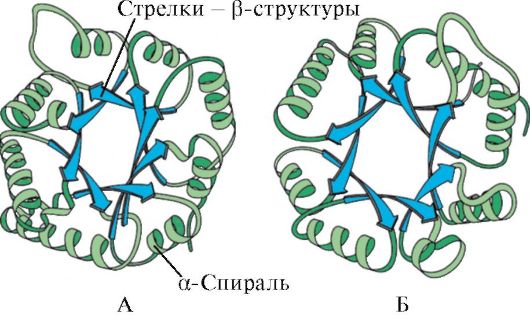 Күріш. 1.6. Триозафосфат изомераза және пируваткиназа доменінің қайталама құрылымы
Күріш. 1.6. Триозафосфат изомераза және пируваткиназа доменінің қайталама құрылымы
 Күріш. 1.7. Иммуноглобулиннің тұрақты доменінің (А) және супероксид дисмутаза ферментінің (В) қайталама құрылымы
Күріш. 1.7. Иммуноглобулиннің тұрақты доменінің (А) және супероксид дисмутаза ферментінің (В) қайталама құрылымы
IN төртінші санатқұрамында қалыпты қайталама құрылымдардың аз мөлшері бар белоктар кіреді. Бұл белоктар құрамында цистеинге бай шағын ақуыздар немесе металлопротеидтер.
Ақуыздың үшінші реттік құрылымы- пептидтік тізбекте бір-бірінен едәуір қашықтықта орналасуы мүмкін аминқышқылдарының радикалдары арасындағы өзара әрекеттесу нәтижесінде түзілетін конформация түрі. Бұл жағдайда белоктардың көпшілігі шар тәрізді (глобулярлы белоктар) кеңістіктік құрылымды құрайды.
Өйткені аминқышқылдарының гидрофобты радикалдары деп аталатындар көмегімен біріктіруге бейім гидрофобты әрекеттесулержәне молекулааралық ван-дер-Ваальс күштерінің әсерінен белок шарының ішінде тығыз гидрофобты өзек түзіледі. Гидрофильді иондалған және иондалмаған радикалдар негізінен белоктың бетінде орналасады және оның суда ерігіштігін анықтайды.
 Күріш. 1.8. Ақуыздың үшінші реттік құрылымының түзілуі кезінде аминқышқылдарының радикалдары арасында пайда болатын байланыс түрлері
Күріш. 1.8. Ақуыздың үшінші реттік құрылымының түзілуі кезінде аминқышқылдарының радикалдары арасында пайда болатын байланыс түрлері
1 - иондық байланыс- оң және теріс зарядталған функционалдық топтар арасында пайда болады;
2 - сутектік байланыс- гидрофильді зарядсыз және кез келген басқа гидрофильді топ арасында кездеседі;
3 - гидрофобты әрекеттесулер- гидрофобты радикалдар арасында пайда болады;
4 - дисульфидтік байланыс- цистеин қалдықтарының SH-топтарының тотығуы және олардың бір-бірімен әрекеттесуі нәтижесінде түзіледі.
Гидрофобты ядродағы гидрофильді аминқышқылдарының қалдықтары бір-бірімен әрекеттесе алады иондықЖәне сутектік байланыстар(1.8-сурет).
Иондық және сутектік байланыстар, сондай-ақ гидрофобты өзара әрекеттесулер әлсіздер қатарына жатады: олардың энергиясы молекулалардың жылулық қозғалысының энергиясынан сәл асып түседі. бөлме температурасы. Ақуыз конформациясы осындай көптеген әлсіз байланыстардың пайда болуымен сақталады. Ақуызды құрайтын атомдар тұрақты қозғалыста болғандықтан, кейбір әлсіз байланыстарды үзіп, басқаларын түзуге болады, бұл полипептидтік тізбектің жеке бөлімдерінің шағын қозғалыстарына әкеледі. Белоктардың кейбірін үзіп, басқа әлсіз байланыстар түзу нәтижесінде конформациясын өзгерту қасиеті деп аталады конформациялық лабильділік.
Адам ағзасында қолдау көрсететін жүйелер бар гомеостаз- сау организм үшін қолайлы белгілі бір шектерде ішкі ортаның тұрақтылығы. Гомеостаз жағдайында конформацияның аздаған өзгерістері бұзылмайды жалпы құрылымжәне белоктардың қызметі. Белоктың функционалдық белсенді конформациясы деп аталады жергілікті конформация.Ішкі ортаның өзгеруі (мысалы, глюкоза, Са иондары, протондар және т.б. концентрациясы) белоктардың конформациясының өзгеруіне және функцияларының бұзылуына әкеледі.
Кейбір белоктардың үшінші реттік құрылымы тұрақтанады дисульфидті байланыстар,екі қалдықтың -SH топтарының әрекеттесуінен түзілген
 Күріш. 1.9. Белок молекуласында дисульфидті байланыстың түзілуі
Күріш. 1.9. Белок молекуласында дисульфидті байланыстың түзілуі
цистеин (1.9-сурет). Жасушаішілік белоктардың көпшілігінде үшінші реттік құрылымында ковалентті дисульфидтік байланыстар болмайды. Олардың болуы жасушадан бөлінетін белоктарға тән, бұл олардың жасушадан тыс жағдайларда үлкен тұрақтылығын қамтамасыз етеді. Сонымен, дисульфидті байланыстар инсулин мен иммуноглобулиндердің молекулаларында болады.
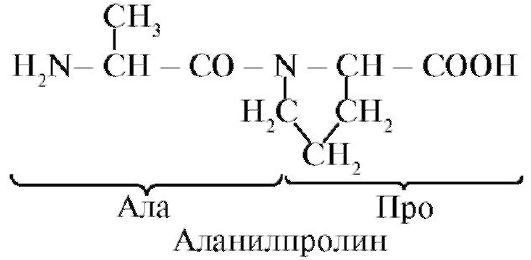
Инсулин- ұйқы безінің β-жасушаларында синтезделетін және қандағы глюкоза концентрациясының жоғарылауына жауап ретінде қанға бөлінетін ақуыздық гормон. Инсулиннің құрылымында полипептидті А- және В-тізбектерін байланыстыратын екі дисульфидтік байланыс және А-тізбектің ішінде бір дисульфидті байланыс бар (1.10-сурет).
 Күріш. 1.10. Инсулин құрылымындағы дисульфидті байланыстар
Күріш. 1.10. Инсулин құрылымындағы дисульфидті байланыстар
5. Белоктардың суперекінші реттік құрылымы.Белоктарда бастапқы құрылымы мен қызметтері әртүрлі, кейде ұқсас комбинациялар және қайталама құрылымдардың интерпозициясы,олар екіншіден жоғары құрылым деп аталады. Ол екінші және үшінші құрылымдар арасында аралық орынды алады, өйткені ол белоктың үшінші реттік құрылымын қалыптастыру кезінде екінші реттік құрылым элементтерінің белгілі бір жиынтығы болып табылады. Суперекіншілік құрылымдардың арнайы атаулары бар, мысалы, «α-спираль-бұрылу-а-спираль», «лейцин сыдырмасы», «мырыш саусақтары» және т.б. Мұндай суперекіншілік құрылымдар ДНҚ-байланыстырушы ақуыздарға тән.
«Лейцин найзағай».Бұл супер қосалқы құрылым екі ақуызды қосу үшін қолданылады. Әрекеттесетін белоктардың бетінде кемінде төрт лейцин қалдығы бар α-спиральді аймақтар бар. α-спиральдағы лейцин қалдықтары бір-бірінен алты аминқышқылдарынан алшақ орналасқан. α-спиральдың әрбір айналымында 3,6 амин қышқылы қалдықтары болғандықтан, әрбір екінші айналымның бетінде лейцин радикалдары кездеседі. Бір ақуыздың α-спиралының лейцин қалдықтары басқа белоктың лейцин қалдықтарымен әрекеттесе алады (гидрофобты әрекеттесу), оларды бір-бірімен байланыстырады (1.11. сурет). Көптеген ДНҚ-байланыстырушы белоктар олигомерлік кешендердің бөлігі ретінде қызмет етеді, мұнда жеке суббірліктер бір-бірімен «лейцин сыдырмалары» арқылы байланысқан.
 Күріш. 1.11. Екі ақуыздың α-спиральді аймақтары арасындағы «лейцин сыдырмасы».
Күріш. 1.11. Екі ақуыздың α-спиральді аймақтары арасындағы «лейцин сыдырмасы».
Гистондар осындай белоктардың мысалы болып табылады. Гистондар- ядролық белоктар, соның ішінде көп саныоң зарядталған аминқышқылдары – аргинин және лизин (80%-ға дейін). Гистон молекулалары осы молекулалардың айтарлықтай омонимдік зарядына қарамастан «лейцинді бекіткіштердің» көмегімен сегіз мономерден тұратын олигомерлік кешендерге біріктіріледі.
«Мырыш саусақ»- ДНҚ-байланыстырушы белоктарға тән суперекіншілік құрылымның нұсқасы белок бетінде ұзартылған фрагмент түрінде болады және құрамында 20-ға жуық амин қышқылы қалдықтары болады (1.12-сурет). «Созылған саусақтың» пішіні төрт амин қышқылы радикалымен - екі цистеин қалдығы және екі гистидин қалдығымен байланысты мырыш атомымен бекітілген. Кейбір жағдайларда гистидин қалдықтарының орнына цистеин қалдықтары болады. Бір-біріне жақын орналасқан екі цистеин қалдығы қалған екі Gisili қалдықтарынан шамамен 12 амин қышқылы қалдықтарынан тұратын Cys тізбегі арқылы бөлінеді. Ақуыздың бұл аймағы α-спиралды құрайды, оның радикалдары ДНҚ негізгі ойығының реттеуші аймақтарымен арнайы байланыса алады. Жеке тұлғаның байланыстыру ерекшелігі
 Күріш. 1.12. «Мырыш саусақ» құрылымын құрайтын ДНҚ-байланыстырушы белоктар бөлімінің бастапқы құрылымы (әріптер осы құрылымды құрайтын аминқышқылдарын көрсетеді)
Күріш. 1.12. «Мырыш саусақ» құрылымын құрайтын ДНҚ-байланыстырушы белоктар бөлімінің бастапқы құрылымы (әріптер осы құрылымды құрайтын аминқышқылдарын көрсетеді)
реттеуші ДНҚ-байланыстырушы протеин «мырыш саусақта» орналасқан аминқышқылдарының қалдықтарының реттілігіне байланысты. Мұндай құрылымдарда, атап айтқанда, транскрипцияны (ДНҚ-дан РНҚ-ға ақпаратты оқу) реттеуге қатысатын стероидты гормондардың рецепторлары бар.
ТАҚЫРЫП 1.2. НЕГІЗДЕРДІҢ ҚЫЗМЕТ КӨРСЕТУІНІҢ НЕГІЗДЕРІ. ПРОТЕИНДІҢ ҚЫЗМЕТІНЕ ӘСЕР ЕТЕТІН ЛИГАНД РЕТІНДЕГІ ДӘРІЛЕР
1. Белоктың активті орталығы және оның лигандпен әрекеттесуі.Үшiншiлiк құрылымның түзiлуi кезiнде функционалдық белсендi белоктың бетiнде, әдетте, ойыстан, бiрiншiлiк құрылымында бiр-бiрiнен алшақ жатқан аминқышқылдарының радикалдары түзетiн орын пайда болады. Белгілі бір ақуыз үшін бірегей құрылымға ие және белгілі бір молекуламен немесе ұқсас молекулалар тобымен арнайы әрекеттесе алатын бұл учаске лигандпен немесе белсенді учаскемен ақуызды байланыстыратын орын деп аталады. Лигандалар - белоктармен әрекеттесетін молекулалар.
Жоғары ерекшелікАқуыздың лигандпен әрекеттесуі активті орталықтың құрылымының лиганд құрылымымен комплементарлы болуымен қамтамасыз етіледі.
толықтыруәрекеттесетін беттердің кеңістіктік және химиялық сәйкестігі болып табылады. Белсенді орталық оның құрамына кіретін лигандаға кеңістіктік сәйкес ғана емес, сонымен қатар белсенді орталыққа кіретін радикалдардың функционалдық топтары мен лиганд арасында лигандты ұстап тұратын байланыстар (иондық, сутегі және гидрофобты әрекеттесулер) түзілуі керек. белсенді орталықта (Cурет 1.13 ).
 Күріш. 1.13. Белоктың лигандпен комплементарлы әрекеттесуі
Күріш. 1.13. Белоктың лигандпен комплементарлы әрекеттесуі
Кейбір лигандтар белоктың белсенді орталығына қосылған кезде белоктардың қызметінде көмекші рөл атқарады. Мұндай лигандтарды кофакторлар, ал құрамында белок емес бөлігі бар белоктар деп аталады. күрделі белоктар(тек протеин бөлігінен тұратын қарапайым белоктардан айырмашылығы). Ақуызға мықтап бекітілген белоксыз бөлік деп аталады протездік топ.Мысалы, миоглобин, гемоглобин және цитохромдардың құрамында белсенді орталыққа мықтап бекітілген протездік топ – құрамында темір ионы бар гем бар. Құрамында гем бар күрделі белоктар гемопротеидтер деп аталады.
Белгілі лигандтар белоктарға қосылғанда бұл белоктардың қызметі көрінеді. Сонымен, қан плазмасындағы ең маңызды ақуыз болып табылатын альбумин белсенді орталыққа май қышқылдары, билирубин, кейбір дәрілік заттар және т.б. сияқты гидрофобты лигандтарды қосу арқылы өзінің тасымалдау қызметін көрсетеді (1.14-сурет).
Пептидтік тізбектің үш өлшемді құрылымымен әрекеттесетін лигандтар төмен молекулалық салмақты органикалық және бейорганикалық молекулалар ғана емес, сонымен қатар макромолекулалар болуы мүмкін:
ДНҚ (жоғарыда ДНҚ байланыстыратын ақуыздармен қарастырылған мысалдар);
полисахаридтер;
 Күріш. 1.14. Генотип пен фенотиптің байланысы
Күріш. 1.14. Генотип пен фенотиптің байланысы
ДНҚ молекуласында кодталған адам белоктарының бірегей бастапқы құрылымы жасушаларда бірегей конформация, белсенді сайт құрылымы және ақуыз функциялары түрінде жүзеге асырылады.
Бұл жағдайларда белок лигандтың байланыстыру орнына сәйкес келетін және оны толықтыратын белгілі бір аймағын таниды. Сонымен, гепатоциттердің бетінде инсулин гормонының рецепторлық белоктары бар, оларда да бар белок құрылымы. Инсулиннің рецептормен әрекеттесуі оның конформациясының өзгеруін және сигналдық жүйелердің белсендірілуін тудырады, тамақтанғаннан кейін гепатоциттерде қоректік заттардың жиналуына әкеледі.
Осылайша, Белоктардың қызметі белоктың белсенді орталығының лигандпен ерекше әрекеттесуіне негізделген.
2. Домен құрылымы және оның белоктардың қызметіндегі рөлі.Глобулярлы белоктардың ұзын полипептидтік тізбектері жиі бірнеше ықшам, салыстырмалы түрде тәуелсіз аймақтарға бүктеледі. Олар глобулярлы белоктарға ұқсайтын тәуелсіз үшінші реттік құрылымға ие және деп аталады домендер.Белоктардың домендік құрылымына байланысты олардың үшінші реттік құрылымын қалыптастыру оңайырақ.
Домен белоктарында лигандтарды байланыстыратын тораптар көбінесе домендер арасында орналасады. Сонымен, трипсин - бұл ұйқы безінің экзокринді бөлігі шығаратын және тағамдық ақуыздарды қорытуға қажет протеолитикалық фермент. Ол екі доменді құрылымға ие және трипсиннің лигандымен - тағамдық ақуызбен байланысу орны екі домен арасындағы ойықта орналасқан. Белсенді орталықта тағамдық ақуыздың белгілі бір учаскесінің тиімді байланысуы және оның пептидтік байланыстарының гидролизі үшін қажетті жағдайлар жасалады.
Белсенді орталық лигандпен әрекеттескенде ақуыздағы әртүрлі домендер бір-біріне қатысты қозғала алады (1.15-сурет).
Гексокиназа- АТФ көмегімен глюкозаның фосфорлануын катализдейтін фермент. Ферменттің белсенді аймағы екі домен арасындағы саңылауда орналасқан. Гексокиназа глюкозамен байланысқан кезде қоршаған домендер жабылады және субстрат ұсталады, онда фосфорлану жүреді (1.15-суретті қараңыз).
 Күріш. 1.15. Гексокиназа домендерінің глюкозамен байланысуы
Күріш. 1.15. Гексокиназа домендерінің глюкозамен байланысуы
Кейбір белоктарда домендер әртүрлі лигандтармен байланысу арқылы тәуелсіз функцияларды орындайды. Мұндай белоктар көп функциялы деп аталады.
3. Дәрілік заттар – белоктардың қызметіне әсер ететін лигандтар.Белоктардың лигандтармен әрекеттесуі ерекше. Дегенмен, ақуыздың конформациялық лабильділігіне және оның белсенді аймағына байланысты, белсенді аймақтағы немесе молекуланың басқа бөлігіндегі ақуызмен де әрекеттесе алатын басқа затты таңдауға болады.
Құрылымы бойынша табиғи лигандқа ұқсас зат деп аталады лигандтың құрылымдық аналогынемесе табиғи емес лиганд. Ол сондай-ақ белсенді аймақтағы ақуызмен әрекеттеседі. Лигандтың құрылымдық аналогы белок қызметін күшейте алады (агонист)және оны азайтыңыз (антагонист).Лиганд және оның құрылымдық аналогтары бір жерде белокпен байланысу үшін бір-бірімен бәсекелеседі. Мұндай заттар деп аталады бәсекеге қабілетті модуляторларбелок функцияларының (регуляторлары). Көптеген препараттар ақуыз ингибиторлары ретінде әрекет етеді. Олардың кейбіреулері табиғи лигандтардың химиялық модификациясы арқылы алынады. Протеин функциясының ингибиторлары есірткі және улану болуы мүмкін.
Атропин М-холинергиялық рецепторлардың бәсекеге қабілетті тежегіші болып табылады.Ацетилхолин - трансмиссиялық нейротрансмиттер жүйке импульсіхолинергиялық синапстар арқылы. Қозуды жүргізу үшін синаптикалық саңылауға бөлінген ацетилхолин постсинапстық мембрананың рецепторы - ақуызбен әрекеттесуі керек. Екі түрі табылды холинергиялық рецепторлар:
М-рецепторацетилхолиннен басқа мускаринмен (шыбынның уыты) селективті әрекеттеседі. М - холинергиялық рецепторлар тегіс бұлшықеттерде болады және ацетилхолинмен әрекеттескенде олардың жиырылуын тудырады;
Н-рецепторникотинмен арнайы байланысады. N-холинергиялық рецепторлар жолақты қаңқа бұлшықеттерінің синапстарында кездеседі.
арнайы ингибитор М-холинергиялық рецепторларатропин болып табылады. Ол белладонна және тауық тұқымдас өсімдіктерде кездеседі.
 Атропиннің құрылымында ацетилхолинге ұқсас функционалды топтары және олардың кеңістіктік орналасуы бар, сондықтан ол М-холинергиялық рецепторлардың бәсекелес тежегіштеріне жатады. Ацетилхолиннің М-холинергиялық рецепторлармен байланысуы тегіс бұлшықеттердің жиырылуын тудыратынын ескере отырып, атропин олардың спазмын жеңілдететін дәрі ретінде қолданылады. (спазмодикалық).Осылайша, көз түбін қарау кезінде көздің бұлшық еттерін босаңсу үшін, сондай-ақ асқазан-ішек коликіндегі спазмды жеңілдету үшін атропинді қолдану белгілі. Орталықта М-холинергиялық рецепторлар да бар жүйке жүйесі(ОЖЖ), сондықтан атропиннің үлкен дозалары орталық жүйке жүйесінің жағымсыз реакциясын тудыруы мүмкін: қозғалтқыш және психикалық қозу, галлюцинация, конвульсия.
Атропиннің құрылымында ацетилхолинге ұқсас функционалды топтары және олардың кеңістіктік орналасуы бар, сондықтан ол М-холинергиялық рецепторлардың бәсекелес тежегіштеріне жатады. Ацетилхолиннің М-холинергиялық рецепторлармен байланысуы тегіс бұлшықеттердің жиырылуын тудыратынын ескере отырып, атропин олардың спазмын жеңілдететін дәрі ретінде қолданылады. (спазмодикалық).Осылайша, көз түбін қарау кезінде көздің бұлшық еттерін босаңсу үшін, сондай-ақ асқазан-ішек коликіндегі спазмды жеңілдету үшін атропинді қолдану белгілі. Орталықта М-холинергиялық рецепторлар да бар жүйке жүйесі(ОЖЖ), сондықтан атропиннің үлкен дозалары орталық жүйке жүйесінің жағымсыз реакциясын тудыруы мүмкін: қозғалтқыш және психикалық қозу, галлюцинация, конвульсия.
Дитилин жүйке-бұлшықет синапстарының қызметін тежейтін Н-холинергиялық рецепторлардың бәсекеге қабілетті агонисі болып табылады.
 Қаңқа бұлшықеттерінің жүйке-бұлшықет синапстарында Н-холинергиялық рецепторлар болады. Олардың ацетилхолинмен әрекеттесуі бұлшықеттердің жиырылуына әкеледі. Кейбір хирургиялық операцияларда, сондай-ақ эндоскопиялық зерттеулерде қаңқа бұлшықеттерінің босаңсуын тудыратын препараттар қолданылады. (бұлшық ет босаңсытқыштары).Оларға ацетилхолиннің құрылымдық аналогы болып табылатын дитилин жатады. Ол Н-холинергиялық рецепторларға қосылады, бірақ ацетилхолинге қарағанда ол ацетилхолинэстераза ферментінің әсерінен өте баяу жойылады. Иондық арналардың ұзақ ашылуы және мембрананың тұрақты деполяризациясы нәтижесінде жүйке импульсінің өткізгіштігі бұзылып, бұлшықет релаксациясы пайда болады. Бастапқыда бұл қасиеттер кураре уында табылды, сондықтан мұндай препараттар деп аталады курариформа.
Қаңқа бұлшықеттерінің жүйке-бұлшықет синапстарында Н-холинергиялық рецепторлар болады. Олардың ацетилхолинмен әрекеттесуі бұлшықеттердің жиырылуына әкеледі. Кейбір хирургиялық операцияларда, сондай-ақ эндоскопиялық зерттеулерде қаңқа бұлшықеттерінің босаңсуын тудыратын препараттар қолданылады. (бұлшық ет босаңсытқыштары).Оларға ацетилхолиннің құрылымдық аналогы болып табылатын дитилин жатады. Ол Н-холинергиялық рецепторларға қосылады, бірақ ацетилхолинге қарағанда ол ацетилхолинэстераза ферментінің әсерінен өте баяу жойылады. Иондық арналардың ұзақ ашылуы және мембрананың тұрақты деполяризациясы нәтижесінде жүйке импульсінің өткізгіштігі бұзылып, бұлшықет релаксациясы пайда болады. Бастапқыда бұл қасиеттер кураре уында табылды, сондықтан мұндай препараттар деп аталады курариформа.
ТАҚЫРЫП 1.3. НЕГІЗДІҢ ДЕНАТУРАЦИЯСЫ ЖӘНЕ ОНЫҢ СОНТАНДЫ ЖАҢА ҚАЛУ МҮМКІНДІГІ
1. Белоктардың табиғи конформациясы әлсіз өзара әрекеттесу, ақуызды қоршап тұрған ортаның құрамы мен қасиеттерінің өзгеруі, химиялық реагенттердің әсері және физикалық факторларолардың конформациясының өзгеруін тудырады (конформациялық лабильдік қасиеті). Көптеген байланыстардың үзілуі нативті конформацияның бұзылуына және ақуыздың денатурациясына әкеледі.
Ақуыздың денатурациясы- бұл белоктың кеңістіктік құрылымын тұрақтандыратын әлсіз байланыстардың үзілуінен туындаған денатурациялаушы агенттердің әсерінен олардың табиғи конформациясының бұзылуы. Денатурация белоктың бірегей үш өлшемді құрылымы мен белсенді орталығының бұзылуымен және оның биологиялық белсенділігінің жоғалуымен бірге жүреді (1.16-сурет).
Бір ақуыздың барлық денатуратталған молекулалары бір белоктың басқа молекулаларынан ерекшеленетін кездейсоқ конформацияға ие болады. Белсенді орталықты құрайтын аминқышқылдарының радикалдары кеңістікте бір-бірінен алшақ болып шығады, яғни. ақуыздың лигандпен арнайы байланысу орны бұзылады. Денатурация кезінде белоктардың бастапқы құрылымы өзгеріссіз қалады.
Биологиялық зерттеулерде және медицинада денатурациялаушы агенттерді қолдану.Биохимиялық зерттеулерде биологиялық материалдағы төмен молекулалық қосылыстарды анықтау алдында әдетте ерітіндіден ақуыздар алынады. Осы мақсатта көбінесе үшхлорсірке қышқылы (TCA) қолданылады. Ерітіндіге TCA қосқаннан кейін денатуратталған ақуыздар тұнбаға түседі және сүзу арқылы оңай жойылады (1.1-кесте).
Медицинада денатурациялаушы заттар көбінесе медициналық аспаптар мен материалдарды автоклавтарда зарарсыздандыру үшін (денатурациялаушы агент - жоғары температура) және антисептиктер ретінде (спирт, фенол, хлорамин) патогенді микрофлорасы бар ластанған беттерді өңдеу үшін қолданылады.
2. Белоктың өздігінен регенерациялануы- белоктардың біріншілік құрылымының, конформациясының және қызметінің детерминизмін дәлелдеу. Жеке белоктар - бұл бір геннің өнімдері, олардың аминқышқылдарының тізбегі бірдей және жасушада бірдей конформацияға ие болады. Белоктың бастапқы құрылымында оның конформациясы мен қызметі туралы ақпарат бар деген іргелі қорытынды кейбір белоктардың (атап айтқанда, рибонуклеаза мен миоглобиннің) өздігінен ренатациялану – денатурациядан кейін өзінің табиғи конформациясын қалпына келтіру қабілеті негізінде жасалды.
Ақуыздың кеңістіктік құрылымдарының қалыптасуы өздігінен құрастыру әдісімен жүзеге асады – бірегей бастапқы құрылымы бар полипептидтік тізбек ерітіндіде ең кіші конформацияны қабылдауға бейім болатын өздігінен жүретін процесс. бос энергия. Денатурациядан кейін бастапқы құрылымын сақтайтын белоктарды регенерациялау қабілеті рибонуклеаза ферментімен жүргізілген тәжірибеде сипатталған.
Рибонуклеаза – РНҚ молекуласындағы жеке нуклеотидтер арасындағы байланысты үзетін фермент. Бұл глобулярлы ақуыздың бір полипептидтік тізбегі бар, оның үшінші реттік құрылымы көптеген әлсіз және төрт дисульфидті байланыстармен тұрақтанған.
Рибонуклеазаны молекуладағы сутектік байланыстарды бұзатын мочевинамен және дисульфидті байланыстарды үзетін тотықсыздандырғышпен өңдеу ферменттің денатурациясына және оның белсенділігінің жоғалуына әкеледі.
Денатурациялаушы агенттерді диализ арқылы жою ақуыздың конформациясы мен функциясын қалпына келтіруге әкеледі, т.б. реанимацияға. (1.17-сурет).
 Күріш. 1.17. Рибонуклеазаның денатурациясы және ренатациялануы
Күріш. 1.17. Рибонуклеазаның денатурациясы және ренатациялануы
А – рибонуклеазаның нативті конформациясы, оның үшінші реттік құрылымында төрт дисульфидтік байланыс бар; В – денатуратталған рибонуклеаза молекуласы;
В – құрылымы мен қызметі қалпына келтірілген ренативтік рибонуклеаза молекуласы
1. 1.2 кестені толтырыңыз.
1.2-кесте. Радикалдардың полярлығына қарай аминқышқылдарының жіктелуі
2. Тетрапептидтің формуласын жаз:
Asp - Pro - Fen - Liz
а) пептидтік магистральді құрайтын пептидтегі қайталанатын топтарды және аминқышқылдарының радикалдарымен ұсынылған айнымалы топтарды бөліп алу;
b) N- және C-терминалдарын белгілеңіз;
в) пептидтік байланыстың астын сызу;
г) бірдей аминқышқылдарынан тұратын басқа пептидті жаз;
д) аминқышқылдарының құрамы ұқсас ықтимал тетрапептидтік нұсқалардың санын санау.
3. Сүтқоректілердің нейрогипофизінің құрылымы жағынан ұқсас және эволюциялық жақын екі пептидті гормоны – окситоцин мен вазопрессинді салыстырмалы талдау мысалында белоктардың біріншілік құрылымының рөлін түсіндіріңіз (1.3-кесте).
1.3-кесте. Окситоцин мен вазопрессиннің құрылысы мен қызметі
 Осыған:
Осыған:
а) екі пептидтің құрамы мен аминқышқылдарының ретін салыстыру;
б) екі пептидтің біріншілік құрылысының ұқсастығын және олардың биологиялық әрекетінің ұқсастығын табу;
в) екі пептидтің құрылысындағы айырмашылықты және олардың қызметтеріндегі айырмашылықты табу;
г) пептидтердің біріншілік құрылымының олардың қызметіне әсері туралы қорытынды жасау.
4. Глобулярлы белоктардың конформациясының қалыптасуының негізгі кезеңдерін сипаттаңыз (екіншілік, үшіншілік құрылымдар, суперекіншілік құрылым туралы түсінік). Белок құрылымдарының түзілуіне қатысатын байланыс түрлерін көрсетіңіз. Қандай аминқышқылдарының радикалдары гидрофобты әрекеттесулердің, иондық, сутектік байланыстардың түзілуіне қатыса алады.
Мысалдар келтір.
5. «Белоктардың конформациялық лабильділігі» ұғымына анықтама беріңіз, оның пайда болу себептері мен маңызын көрсетіңіз.
6. Келесі сөз тіркесінің мағынасын түсіндіріңіз: «Белоктар лигандпен ерекше әрекеттесуіне негізделген» терминдерді қолданып, олардың мағынасын түсіндіріңіз: ақуыз конформациясы, белсенді орын, лиганд, комплементарлық, ақуыз қызметі.
7. Мысалдардың бірін пайдаланып, домендердің не екенін және олардың белоктардың қызметіндегі рөлі қандай екенін түсіндіріңіз.
ӨЗІН-ӨЗІ БАҚЫЛАУ Тапсырмалары
1. Сәйкестік орнату.
Амин қышқылы радикалындағы функционалдық топ:
A. Карбоксил тобы B. Гидроксил тобы C. Гуанидин тобы D. Тиол тобы E. Аминотобы
2. Дұрыс жауаптарды таңдаңыз.
Полярлы зарядсыз радикалдары бар аминқышқылдары:
A. Tsis B. Asn
B. Glu G. Үш
3. Дұрыс жауаптарды таңдаңыз.
Амин қышқылының радикалдары:
A. Бастапқы құрылымның ерекшелігін қамтамасыз ету B. Үшіншілік құрылымның қалыптасуына қатысу
B. Ақуыздың бетінде орналасқандықтан, оның ерігіштігіне әсер етеді D. Белсенді орталық құрайды
D. Пептидтік байланыстың түзілуіне қатысу
4. Дұрыс жауаптарды таңдаңыз.
Аминқышқылдарының радикалдары арасында гидрофобты әрекеттесулер пайда болуы мүмкін:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Дұрыс жауаптарды таңдаңыз.
Аминқышқылдарының радикалдары арасында иондық байланыс түзілуі мүмкін:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Goese Asp D. Asn Сәуір
6. Дұрыс жауаптарды таңдаңыз.
Аминқышқылдарының радикалдары арасында сутектік байланыс түзілуі мүмкін:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Сәйкестік орнату.
Белок құрылымының түзілуіне қатысатын байланыс түрі:
A. Бастапқы құрылым B. Қосалқы құрылым
B. Үшіншілік құрылым
D. Жоғары екіншілік құрылым E. Конформация.
1. Пептидтік магистраль атомдары арасындағы сутектік байланыс
2. Амин қышқылы радикалдарының функционалдық топтары арасындағы әлсіз байланыстар
3. Амин қышқылдарының α-амин және α-карбоксил топтары арасындағы байланыс
8. Дұрыс жауаптарды таңдаңыз. Трипсин:
A. Протеолитикалық фермент B. Құрамында екі домен бар
B. Крахмалды гидролиздейді
D. Белсенді орталық домендер арасында орналасқан. D. Екі полипептидтік тізбектен тұрады.
9. Дұрыс жауаптарды таңдаңыз. Атропин:
A. Нейротрансмиттер
B. Ацетилхолиннің құрылымдық аналогы
B. Н-холинергиялық рецепторлармен әрекеттеседі
G. Холинергиялық синапс арқылы жүйке импульсінің өткізілуін күшейтеді
D. М-холинергиялық рецепторлардың бәсекелес ингибиторы
10. Дұрыс мәлімдемелерді таңдаңыз. Белоктарда:
A. Бастапқы құрылым өзінің белсенді сайтының құрылымы туралы ақпаратты қамтиды
B. Белсенді орталық бастапқы құрылым деңгейінде қалыптасады
B. Конформация коваленттік байланыстармен қатаң бекітілген
D. Белсенді учаске ұқсас лигандтар тобымен әрекеттесе алады
белоктардың конформациялық лабильділігіне байланысты D. Өзгеруі қоршаған орта, белсендінің жақындығына әсер етуі мүмкін
орталықтан лигандқа
1. 1-C, 2-D, 3-B.
3. A, B, C, D.
7. 1-В, 2-Д, 3-А.
8. A, B, C, D.
НЕГІЗГІ ТЕРМИНДЕР МЕН ТҮСІНІКТЕР
1. Белок, полипептид, амин қышқылдары
2. Біріншілік, екіншілік, үшіншілік белок құрылымдары
3. Конформация, нативті белок конформациясы
4. Белоктағы коваленттік және әлсіз байланыстар
5. Конформациялық лабильділік
6. Белсенді протеин
7. Лигандалар
8. Белоктардың қатпарлануы
9. Лигандтардың құрылымдық аналогтары
10. Домен белоктары
11. Қарапайым және күрделі белоктар
12. Ақуызды денатурациялау, денатурациялаушы заттар
13. Ақуыз регенерациясы
Мәселелерді шешу
«Белоктардың құрылымдық ұйымдастырылуы және олардың қызмет ету негіздері»
1. Белоктың негізгі қызметі – гемоглобин А (HbA) – оттегін ұлпаларға тасымалдау. адам популяциясына белгілі көпше түрлеріБұл ақуыздың қасиеттері мен қызметі өзгерген – анормальды гемоглобиндер деп аталады. Мысалы, орақ жасушалы анемия (HbS) бар науқастардың эритроциттерінен табылған гемоглобин S оттегінің парциалды қысымы төмен болған жағдайда (веноздық қанда болатындай) ерігіштігі төмен екендігі анықталды. Бұл осы ақуыздың агрегаттарының пайда болуына әкеледі. Ақуыз өз қызметін жоғалтады, тұнбаға түседі, эритроциттер жиналады дұрыс емес пішін(олардың кейбіреулері орақ пішінді құрайды) және көкбауырда әдеттегіден тезірек жойылады. Нәтижесінде орақ жасушалы анемия дамиды.
HvA-ның бастапқы құрылымындағы жалғыз айырмашылық гемоглобиннің β-тізбегінің N-терминал аймағында табылды. β-тізбектің N-соңғы аймақтарын салыстырыңыз және белоктың біріншілік құрылымындағы өзгерістер оның қасиеттері мен қызметіне қалай әсер ететінін көрсетіңіз.
 Осыған:
Осыған:
а) HvA қандай аминқышқылдарының формулаларын жазыңыз және осы аминқышқылдарының қасиеттерін салыстырыңыз (полярлық, заряд).
б) ұлпада ерігіштіктің төмендеуі және оттегінің тасымалдануының бұзылуының себебі туралы қорытынды жасау.
2. Суретте лигандтарды байланыстыру орталығы (активті орталық) бар ақуыздың құрылымының диаграммасы көрсетілген. Лиганды таңдауда белок неліктен селективті болатынын түсіндіріңіз. Осыған:
а) белоктың активті орталығы қандай екенін есте сақтаңыз және суретте көрсетілген белоктың белсенді орталығының құрылымын қарастырыңыз;
б) белсенді орталықты құрайтын аминқышқылдарының радикалдарының формулаларын жазу;
в) ақуыздың белсенді аймағымен арнайы әрекеттесе алатын лигандты сызыңыз. Онда белсенді орталықты құрайтын аминқышқылдарының радикалдарымен байланыс түзуге қабілетті функционалдық топтарын көрсетіңіз;
г) активті орталықтың лиганд пен аминқышқылдарының радикалдары арасында пайда болатын байланыс түрлерін көрсетіңіз;
д) Белоктың лигандпен әрекеттесу ерекшелігінің негізін түсіндіріңіз.
 3.
Суретте ақуыздың және бірнеше лигандтардың белсенді орны көрсетілген.
3.
Суретте ақуыздың және бірнеше лигандтардың белсенді орны көрсетілген.
Лигандтардың қайсысы белоктың белсенді жерімен әрекеттесуі мүмкін екенін және неліктен екенін анықтаңыз.
 Белок-лиганд кешенінің түзілуі кезінде қандай байланыс түрлері пайда болады?
Белок-лиганд кешенінің түзілуі кезінде қандай байланыс түрлері пайда болады?
4. Табиғи ақуыз лигандтарының құрылымдық аналогтары белоктардың белсенділігін өзгерту үшін дәрілік заттар ретінде қолданылуы мүмкін.
Ацетилхолин жүйке-бұлшықет синапстарында қозуды тасымалдау медиаторы болып табылады. Ацетилхолин белоктармен – қаңқа бұлшықеттерінің постсинапстық мембранасының рецепторларымен әрекеттескенде иондық арналар ашылып, бұлшықеттің жиырылуы пайда болады. Дитилин - бұлшық еттерді босаңсу үшін кейбір операцияларда қолданылатын препарат, өйткені ол жүйке импульстарының жүйке-бұлшықет синапстары арқылы берілуін бұзады. Дитилиннің бұлшықет босаңсытқышы ретінде әсер ету механизмін түсіндіріңіз. Осыған:
а) ацетилхолин мен дитилиннің формулаларын жазып, құрылысын салыстыру;
б) дитилиннің босаңсыту әсерінің механизмін сипаттаңыз.
5. Кейбір ауруларда науқастың дене температурасы көтеріледі, бұл дененің қорғаныс реакциясы ретінде қарастырылады. Дегенмен, жоғары температура дененің ақуыздарына зиян келтіреді. Неліктен 40 °C жоғары температурада белоктардың қызметі бұзылып, адам өміріне қауіп төнетінін түсіндіріңіз. Мұны істеу үшін есте сақтаңыз:
1) Белоктардың құрылымы және оның құрылымын нативті конформацияда ұстайтын байланыстар;
2) Температура жоғарылағанда белоктардың құрылысы мен қызметі қалай өзгереді?;
3) Гомеостаз дегеніміз не және адам денсаулығын сақтау неліктен маңызды.
Модульдік блок 2 РЕГАТОРЛЫҚ ӘСЕР ҮШІН МАҚСАТТАР РЕТІНДЕ ОЛИГОМЕРЛІК БЕЛГІЛЕР. НЕГІЗДІҢ ҚҰРЫЛЫМДЫҚ ЖӘНЕ ФУНКЦИЯЛЫҚ ТҮРЛІЛІГІ. БЕЛГІЛЕРДІ БӨЛУ ЖӘНЕ ТАЗАЛАУ ӘДІСТЕРІ
Оқу мақсаттары Істей білу:
1.Олигомерлі белоктардың құрылысы мен қызметтерінің ерекшеліктері туралы білімдерін олардың қызметтерін реттеудің адаптивті механизмдерін түсіну үшін пайдалану.
2. Шаперондардың жасушадағы ақуыз конформациясының синтезі мен сақталуындағы рөлін түсіндіріңіз.
3. Организмде синтезделетін белоктардың құрылымдары мен қызметтерінің алуан түрлілігі арқылы тіршілік көріністерінің әртүрлілігін түсіндіру.
4. Туысқан гемопротеидтерді – миоглобин мен гемоглобинді, сондай-ақ иммуноглобулиндердің бес класы белоктарының өкілдерін салыстыра отырып, белоктардың құрылысы мен олардың қызметі арасындағы байланысты талдаңыз.
5. Белоктардың физика-химиялық қасиеттерінің ерекшеліктері туралы білімдерін оларды басқа белоктар мен қоспалардан тазарту әдістерін таңдау үшін қолдану.
6. Клиникалық диагнозды растау немесе нақтылау үшін қан плазмасының ақуыздарының сандық және сапалық құрамының нәтижелерін интерпретациялау.
Білу:
1. Гемоглобин мысалында олигомерлі белоктардың құрылыс ерекшеліктері және олардың қызметтерін реттеудің адаптивті механизмдері.
2. Шаперондардың құрылысы мен қызметтері және олардың жасушадағы белоктардың табиғи конформациясын сақтаудағы маңызы.
3. Иммуноглобулиндер мысалында белоктарды конформациясы мен қызметтерінің ұқсастығына қарай тұқымдастарға топтастыру принциптері.
4. Белоктарды олардың физика-химиялық қасиеттерінің ерекшеліктеріне қарай бөлу әдістері.
5. Қан плазмасының электрофорезі белоктардың сапалық және сандық құрамын бағалау әдісі ретінде.
ТАҚЫРЫП 1.4. ГЕМОГЛОБИН МЫСАЛЫНДА ОЛИГОМЕРЛІК НЕГІЗДЕРДІҢ ҚҰРЫЛЫСЫ МЕН ҚЫЗМЕТІНІҢ ЕРЕКШЕЛІКТЕРІ
1. Көптеген ақуыздарда бірнеше полипептидтік тізбектер бар. Мұндай белоктар деп аталады олигомерлік,және жеке тізбектер протомерлер.Олигомерлі белоктардағы протомерлер көптеген әлсіз ковалентті емес байланыстармен (гидрофобты, иондық, сутегі) байланысады. Өзара әрекеттесу
протомерлердің арқасында жүзеге асырылады толықтыруолардың жанасу беттері.
Олигомерлі белоктардағы протомерлердің саны әр түрлі болуы мүмкін: гемоглобиннің құрамында 4 протомер, аспартатаминотрансфераза ферментінде - 12 протомер, ал темекі мозаикалық вирусының ақуызында ковалентті емес байланыстар арқылы байланысқан 2120 протомер бар. Сондықтан олигомерлі белоктар өте жоғары молекулалық салмаққа ие болуы мүмкін.
Бір протомердің басқалармен әрекеттесуін белоктың лигандпен әрекеттесуінің ерекше жағдайы ретінде қарастыруға болады, өйткені әрбір протомер басқа протомерлер үшін лиганд қызметін атқарады. Белоктағы протомерлердің саны мен қосылу әдісі деп аталады төрттік белок құрылымы.
Белоктардың құрамында бірдей немесе әртүрлі құрылымның протомерлері болуы мүмкін, мысалы, гомодимерлер – екі бірдей протомерлері бар белоктар, ал гетеродимерлер – екі түрлі протомерлері бар белоктар.
Егер белоктарда әртүрлі протомерлер болса, онда оларда құрылымы жағынан әр түрлі лигандтары бар байланыстыру орталықтары пайда болуы мүмкін. Лиганд белсенді орталықпен байланысқанда бұл ақуыздың қызметі көрінеді. Басқа протомерде орналасқан орталық аллостериялық (белсендіден басқа) деп аталады. Байланыс аллостериялық лиганд немесе эффектор,ол реттеуші функцияны орындайды (1.18-сурет). Аллостериялық орталықтың эффектормен әрекеттесуі конформациялық лабильділігіне байланысты бүкіл олигомерлі белоктың құрылымында конформациялық өзгерістерді тудырады. Бұл белсенді аймақтың белгілі бір лигандқа жақындығына әсер етеді және осы ақуыздың қызметін реттейді. Олигомерлі белоктың кем дегенде бір лигандпен әрекеттесуі кезінде барлық протомерлердің конформациясы мен қызметінің өзгеруін кооперативтік конформация өзгерісі деп атайды. Белок қызметін күшейтетін эффекторлар деп аталады активаторларжәне оның қызметін төмендететін эффекторлар - ингибиторлар.
Сонымен, олигомерлі белоктарда, сондай-ақ домендік құрылымы бар белоктарда мономерлі белоктармен салыстырғанда жаңа қасиет пайда болады – функцияларды аллостериялық реттеу қабілеті (ақуызға әртүрлі лигандтарды қосу арқылы реттеу). Мұны миоглобин мен гемоглобиннің бір-бірімен тығыз байланысты екі күрделі белоктардың құрылымдары мен қызметтерін салыстыру арқылы көруге болады.
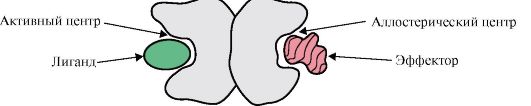 Күріш. 1.18. Димерлі белоктың құрылымының диаграммасы
Күріш. 1.18. Димерлі белоктың құрылымының диаграммасы
2. Кеңістіктік құрылымдардың қалыптасуы және миоглобиннің қызметі.
Миоглобин (Мб) - қызыл бұлшықеттерде кездесетін ақуыз, оның негізгі қызметі бұлшықеттердің қарқынды жұмысына қажетті O 2 қорын жасау болып табылады. МБ – құрамында белок бөлігі – apoMB және белоксыз бөлігі – гем бар күрделі ақуыз. apoMB-ның біріншілік құрылымы оның ықшам глобулярлық конформациясын және миоглобиннің белоксыз бөлігі гем бекітілген белсенді орталықтың құрылымын анықтайды. Қаннан бұлшық еттерге оттегі миоглобиннің құрамындағы Fe+2 гемімен байланысады. МБ – мономерлік ақуыз, O 2-ге өте жоғары жақындығы бар, сондықтан миоглобинмен оттегі тек қарқынды бұлшықет жұмысы кезінде, О 2 парциалды қысымы күрт төмендегенде бөлінеді.
МБ конформациясының қалыптасуы.Қызыл бұлшықеттерде, рибосомаларда трансляция кезінде 153 амин қышқылы қалдықтарының белгілі бір тізбегімен ұсынылған МБ-ның бастапқы құрылымының синтезі жүреді. Mv-нің екінші реттік құрылымы А-дан Н-ге дейінгі латын әріптері деп аталатын сегіз α-спиральдан тұрады, олардың арасында спиральданбаған бөлімдер бар. Mv-тің үшінші реттік құрылымы ықшам шар тәрізді пішінге ие, оның ойығында F және E α-спиральдар арасында белсенді орталық орналасқан (1.19-сурет).
 Күріш. 1.19. Миоглобиннің құрылысы
Күріш. 1.19. Миоглобиннің құрылысы
3. МВ белсенді орталығының құрылымы мен қызмет ету ерекшеліктері. Mv активті орталығы негізінен біріншілік құрылымы бойынша бір-бірінен алшақ орналасқан гидрофобты аминқышқылдарының радикалдарынан түзіледі (мысалы, Tri 3 9 және Phen 138) Суда нашар еритін лигандтар, гем және O 2, белсенді орталыққа бекітілген. Гем – спецификалық apoMv лиганд (1.20-сурет), оның негізінде метенил көпірлері арқылы қосылған төрт пиррол сақиналары жатыр; ортасында төрт координациялық байланыс арқылы пиррол сақиналарының азот атомдарымен байланысқан Fe+ 2 атомы бар. Мв белсенді орталығында аминқышқылдарының гидрофобты радикалдарынан басқа гидрофильді радикалдары бар екі аминқышқылының қалдықтары да бар - Gis E 7(Gis 64) және Gis F 8(Оның 93) (1.21-сурет).
 Күріш. 1.20. Гемнің құрылымы – миоглобин мен гемоглобиннің белоксыз бөлігі
Күріш. 1.20. Гемнің құрылымы – миоглобин мен гемоглобиннің белоксыз бөлігі
 Күріш. 1.21. Апомиоглобин мен гемоглобин протомерлерінің белсенді аймағында гем мен О 2 орналасуы
Күріш. 1.21. Апомиоглобин мен гемоглобин протомерлерінің белсенді аймағында гем мен О 2 орналасуы
Гем оның F 8-мен темір атомы арқылы ковалентті байланысқан. О 2 гем жазықтығының екінші жағындағы темірге бекітіледі. Оның E 7 O 2 дұрыс бағытталуы үшін қажет және Fe + 2 гемге оттегінің қосылуын жеңілдетеді.
Gis F 8 Fe+ 2-мен координациялық байланыс түзеді және гемді белсенді жерде берік бекітеді. Gis E 7 Fe+2 геммен әрекеттесу кезінде басқа лиганд – О 2 активті орталығында дұрыс бағытталуы үшін қажет. Гемдік микроорта O 2-нің Fe + 2-мен күшті, бірақ қайтымды байланысуы үшін жағдай жасайды және судың гидрофобты белсенді орталыққа түсуіне жол бермейді, бұл оның Fe + 3 -ке дейін тотығуына әкелуі мүмкін.
МБ мономерлік құрылымы және оның белсенді орталығы ақуыздың O 2-ге жоғары жақындығын анықтайды.
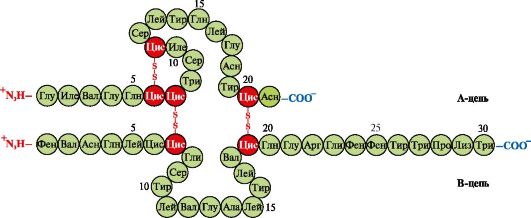
4. Hb-ның олигомерлік құрылымы және лигандтар арқылы O 2-ге Hb сәйкестігінің реттелуі. Адамның гемоглобиндері- белоктар тұқымдасы, сонымен қатар күрделі белоктарға (гемопротеидтерге) жататын миоглобин. Олардың тетрамерлі құрылымы бар және екі α-тізбекті қамтиды, бірақ басқа екі полипептидтік тізбектердің (2α-, 2х-тізбектері) құрылымы бойынша ерекшеленеді. Екінші полипептидтік тізбектің құрылымы Hb осы формаларының қызмет ету ерекшеліктерін анықтайды. Ересек эритроциттердегі гемоглобиннің шамамен 98% құрайды гемоглобин А(2α-, 2p-тізбектері).
Ұрықтың дамуы кезінде гемоглобиндердің екі негізгі түрі бар: эмбриональды HB(2α, 2ε), ол ұрық дамуының ерте кезеңдерінде кездеседі және гемоглобин F (ұрық)- (2α, 2γ), ол ұрық дамуының алтыншы айында ерте ұрықтың гемоглобинін ауыстырады және тек туғаннан кейін Hb A ауыстырылады.
Hv A – ересек эритроциттерде кездесетін миоглобинге (Mv) жататын ақуыз. Оның жеке протомерлерінің құрылымы миоглобиндікіне ұқсас. Миоглобин мен гемоглобин протомерлерінің қайталама және үшінші құрылымдары олардың полипептидтік тізбектерінің біріншілік құрылымында тек 24 аминқышқылдарының қалдықтары бірдей болғанына қарамастан өте ұқсас (гемоглобин протомерлерінің екінші реттік құрылымында, миоглобин сияқты, сегіз α-спираль, А-дан Н-ге дейінгі латын әріптерімен белгіленеді және үшінші құрылым ықшам глобула түрінде болады). Бірақ миоглобиннен айырмашылығы, гемоглобин олигомерлік құрылымға ие, ковалентті емес байланыстармен қосылған төрт полипептидтік тізбектен тұрады (1.22-сурет).
Әрбір Hb протомері белокты емес бөлікпен – геммен және көршілес протомерлермен байланысты. Нв-ның белоктық бөлігінің геммен байланысы миоглобиндікіне ұқсас: белоктың белсенді орталығында гемнің гидрофобты бөліктері His F 8 және His E 7-ден басқа гидрофобты аминқышқылдарының радикалдарымен қоршалған. , олар гем жазықтығының екі жағында орналасқан және ақуыздың қызметінде және оның оттегімен байланысуында ұқсас рөл атқарады (миоглобиннің құрылымын қараңыз).
 Күріш. 1.22. Гемоглобиннің олигомерлік құрылымы
Күріш. 1.22. Гемоглобиннің олигомерлік құрылымы
Сонымен қатар, Gis E 7маңызды орындайды қосымша рөл NV қызметінде. Бос гемнің СО-ға жақындығы O 2-ге қарағанда 25000 есе жоғары. СО организмде аз мөлшерде түзіледі және оның гемге жоғары жақындығын ескере отырып, ол жасуша тіршілігіне қажетті O 2 тасымалдануын бұзуы мүмкін. Алайда гемоглобиннің құрамында гемнің көміртегі тотығына жақындығы His-тің белсенді орталығында Е 7 болуына байланысты O 2-ге жақындығы 200 есе ғана асып түседі. Бұл амин қышқылының қалдығы гемнің О2-мен байланысуына оңтайлы жағдай жасайды және гемнің СО-мен әрекеттесуін әлсіретеді.
5. Hb негізгі қызметі – өкпеден тіндерге O 2 тасымалдау. O 2-ге өте жоғары жақындығы бар және қызыл бұлшықеттерде оттегін сақтау қызметін атқаратын мономерлі миоглобиннен айырмашылығы, гемоглобиннің олигомерлік құрылымы мыналарды қамтамасыз етеді:
1) өкпедегі Hb оттегімен тез қанығуы;
2) Hb салыстырмалы жоғары парциалды қысым O 2 (20-40 мм рт.ст.) кезінде тіндерге оттегін шығару қабілеті;
3) Hb-ның O 2-ге жақындығын реттеу мүмкіндігі.
6. Гемоглобин протомерлерінің конформациясындағы кооперативтік өзгерістер өкпедегі O 2 байланысын және оның тіндерге оралуын тездетеді. Өкпеде O2 жоғары парциалды қысымы оның төрт протомердің (2α және 2β) белсенді орнында Hb-мен байланысуына ықпал етеді. Әрбір протомердің белсенді орталығы, миоглобиндегідей, гидрофобты қалтадағы екі α-спираль (F және E) арасында орналасқан. Оның құрамында белокты емес бөлік – гем бар, белок бөлігіне көптеген әлсіз гидрофобты әрекеттесулер және Fe 2 + гем мен Оның F 8 арасындағы бір күшті байланыс (1.21-суретті қараңыз).
Дезоксигемоглобинде оның F 8-мен осы байланысына байланысты Fe 2 + атомы гем жазықтығынан гистидинге қарай шығып тұрады. O 2-нің Fe 2+-мен байланысуы бір бос координациялық байланыстың көмегімен His E 7 аймағындағы гемнің екінші жағында жүреді. Оның E 7 гемдік темірмен O 2 байланысуы үшін оңтайлы жағдайларды қамтамасыз етеді.
Бір протомердің Fe +2 атомына O 2 қосылуы оның гем жазықтығына, ал оның артында онымен байланысты гистидин қалдығына жылжиды.
 Күріш. 1.23. Гемоглобин протомерінің O 2-мен біріктірілген конформациясының өзгеруі
Күріш. 1.23. Гемоглобин протомерінің O 2-мен біріктірілген конформациясының өзгеруі
Бұл барлық полипептидтік тізбектердің конформациялық лабильділігіне байланысты конформациясының өзгеруіне әкеледі. Басқа тізбектердің конформациясын өзгерту олардың келесі O 2 молекулаларымен әрекеттесуін жеңілдетеді.
Төртінші O 2 молекуласы гемоглобинге біріншіге қарағанда 300 есе оңай қосылады (1.24-сурет).
 Күріш. 1.24. Гемоглобин протомерлерінің О 2-мен әрекеттесуі кезінде конформациясының кооперативтік өзгерістері
Күріш. 1.24. Гемоглобин протомерлерінің О 2-мен әрекеттесуі кезінде конформациясының кооперативтік өзгерістері
Тіндерде әрбір келесі O 2 молекуласы алдыңғысына қарағанда оңай бөлінеді, сонымен қатар протомер конформациясындағы бірлескен өзгерістерге байланысты.
7. Катаболизм кезінде түзілетін СО 2 және Н+ органикалық заттар, гемоглобиннің O 2-ге жақындығын олардың концентрациясына пропорционалды түрде төмендетеді. Жасушаның жұмыс істеуіне қажетті энергия негізінен гемоглобин арқылы өкпеден жеткізілетін O 2 көмегімен органикалық заттардың тотығуы кезінде митохондрияларда өндіріледі. Органикалық заттардың тотығуы нәтижесінде соңғы өнімдеролардың ыдырауы: CO 2 және K 2 O, олардың мөлшері жүріп жатқан тотығу процестерінің қарқындылығына пропорционалды.
СО 2 жасушалардан қанға таралады және эритроциттерге енеді, онда карбангидраза ферментінің әсерінен ол қанға айналады. көмір қышқылы. Бұл әлсіз қышқыл протонға және бикарбонат ионына диссоциацияланады.
H+ GIS радикалдарына қосылуға қабілетті 14 6 гемоглобиннің α- және β-тізбектерінде, яғни. гемнен алыс жерлерде. Гемоглобиннің протондануы оның O 2-ге жақындығын төмендетеді, oxyHb-ден O 2-нің жойылуына, deoxyHb түзілуіне ықпал етеді және түзілген протондар санына пропорционалды түрде тіндердің оттегімен қамтамасыз етілуін арттырады (1.25-сурет).
Эритроциттердегі Н+ концентрациясының артуына байланысты бөлінетін оттегі мөлшерінің артуы Бор эффектісі деп аталады (бұл әсерді алғаш ашқан дат физиологы Кристиан Бордың атымен).
Өкпеде оттегінің жоғары парциалды қысымы оның deoxyHb-мен байланысуына ықпал етеді, бұл ақуыздың H+-ға жақындығын төмендетеді. Карбангидраза әсерінен бөлінген протондар бикарбонаттармен әрекеттесіп, CO 2 және H 2 O түзеді.
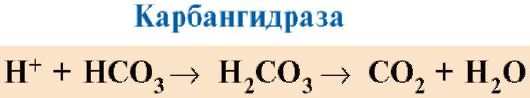
 Күріш. 1.25. Hb-ның O 2-ге жақындығының CO 2 және протондар концентрациясына тәуелділігі (Бор эффектісі):
Күріш. 1.25. Hb-ның O 2-ге жақындығының CO 2 және протондар концентрациясына тәуелділігі (Бор эффектісі):
А- СО 2 және Н+ концентрациясының Hb бар кешеннен O 2 бөлінуіне әсері (Бор эффектісі); Б- өкпедегі дезоксигемоглобинді оттегімен қамтамасыз ету, СО 2 түзілуі және бөлінуі.
Алынған СО 2 альвеолярлы кеңістікке еніп, дем шығарылған ауамен бірге шығарылады. Осылайша, тіндерде гемоглобин бөлетін оттегінің мөлшері органикалық заттардың катаболизмі өнімдерімен реттеледі: заттардың ыдырауы неғұрлым қарқынды болса, мысалы, физикалық жүктеме кезінде, СО 2 және H + концентрациясы соғұрлым жоғары болады және соғұрлым көп болады. тіндер оттегіні H-ның O 2-ге жақындығының төмендеуі нәтижесінде алады.
8. Лиганды – 2,3-бисфосфоглицерат арқылы O 2-ге Hb жақындығын аллостериялық реттеу.Эритроциттерде гемоглобиннің аллостериялық лиганды 2,3-бисфосфоглицерат (2,3-БПГ) глюкозаның тотығу өнімінен – 1,3-бифосфоглицераттан синтезделеді. Қалыпты жағдайда 2,3-BPG концентрациясы жоғары және Hb концентрациясымен салыстырмалы. 2,3-BPG күшті теріс заряды -5.
 Тіндік капиллярлардағы бисфосфоглицерат дезоксигемоглобинмен байланысу арқылы тіндердегі оттегінің шығуын арттырады, Hb-ның O 2-ге жақындығын төмендетеді.
Тіндік капиллярлардағы бисфосфоглицерат дезоксигемоглобинмен байланысу арқылы тіндердегі оттегінің шығуын арттырады, Hb-ның O 2-ге жақындығын төмендетеді.
Тетрамерлі гемоглобин молекуласының ортасында қуыс бар. Ол барлық төрт протомердің аминқышқылдарының қалдықтарынан түзіледі (1.22-суретті қараңыз). Тіндік капиллярларда Hb протонациясы (Бор эффектісі) темір гем мен O 2 арасындағы байланысты үзеді. Молекулада
дезоксигемоглобин, оксигемоглобинмен салыстырғанда, протомерлерді байланыстыратын қосымша иондық байланыстар пайда болады, нәтижесінде орталық қуыстың мөлшері оксигемоглобинмен салыстырғанда артады. Орталық қуыс 2,3-БПГ-ның гемоглобинге қосылу орны болып табылады. Орталық қуыстың өлшемдерінің айырмашылығына байланысты 2,3-БПГ тек дезоксигемоглобинге қосыла алады.
2,3-BPG гемоглобинмен ақуыздың белсенді орындарынан қашықтағы аймақта әрекеттеседі және аллостериялық(реттеуші) лигандтар, ал орталық қуысы Hb болып табылады аллостериялық орталық. 2,3-BPG күшті теріс зарядқа ие және екі Hb β-тізбегінің бес оң зарядталған тобымен әрекеттеседі: N-терминал α-амин тобы Val және Lys 82 Gis 143 радикалдары (1.26-сурет).
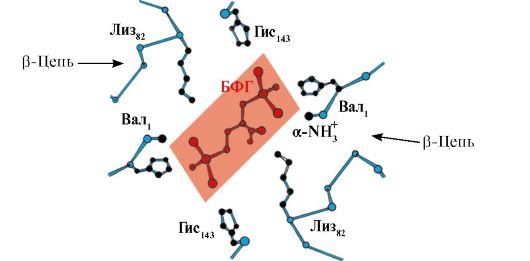 Күріш. 1.26. Дезоксигемоглобиннің орталық қуысында BPG
Күріш. 1.26. Дезоксигемоглобиннің орталық қуысында BPG
BPG әрбір β-тізбекте үш оң зарядталған топтармен байланысады.
Ұлпа капиллярларында пайда болған дезоксигемоглобин 2,3-БПГ-мен әрекеттеседі, β-тізбектердің оң зарядты радикалдары мен теріс зарядталған лиганд арасында иондық байланыстар түзіледі, олар белок конформациясын өзгертеді және Hb-ның O 2-ге жақындығын төмендетеді. Hb-ның O 2-ге жақындығының төмендеуі O 2-нің тінге тиімдірек бөлінуіне ықпал етеді.
Өкпеде жоғары парциалды қысымда оттегі Hb-мен әрекеттеседі, темір геміне қосылады; бұл жағдайда белоктың конформациясы өзгереді, орталық қуыс төмендейді, ал 2,3-БПГ аллостериялық орталықтан ығысады.
Осылайша, олигомерлі белоктар мономерлі белоктармен салыстырғанда жаңа қасиеттерге ие. Лигандтардың учаскелерде қосылуы,
бір-бірінен кеңістіктік алыс (аллостериялық), бүкіл ақуыз молекуласында конформациялық өзгерістерді тудыруға қабілетті. Реттеуші лигандтармен әрекеттесуіне байланысты конформация өзгереді және белок молекуласының қызметі қоршаған орта өзгерістеріне бейімделеді.
ТАҚЫРЫП 1.5. ЖАСУША ЖАҒДАЙЫНДАҒЫ БЕЛГІЛІКТЕРДІҢ НАТИВИ КОНФОРМАЦИЯСЫН САҚТАУ
Жасушаларда полипептидтік тізбектердің синтезі кезінде олардың мембраналар арқылы жасушаның сәйкес бөлімдеріне тасымалдануы, қатпарлану процесінде (нативті конформацияның түзілуі) және олигомерлі белоктардың жиналуы кезінде, сонымен қатар олардың жұмыс істеуі кезінде, аралық , белок құрылымында агрегацияға бейім, тұрақсыз конформациялар пайда болады. Әдетте белок молекуласының ішінде табиғи конформациясында жасырылған гидрофобты радикалдар бетінде тұрақсыз конформацияда пайда болады және суда бірдей нашар еритін басқа ақуыздар топтарымен қосылуға бейім. Барлық белгілі организмдердің жасушаларында жасуша белоктарының оңтайлы қатпарлануын қамтамасыз ететін, қызмет ету кезінде олардың нативті конформациясын тұрақтандыратын, ең бастысы гомеостаз бұзылған жағдайда жасушаішілік белоктардың құрылымы мен қызметін сақтайтын арнайы белоктар табылды. Бұл белоктар деп аталады «каперондар»француз тілінен аударғанда «күту» дегенді білдіреді.
1. Молекулярлық шаперондар және олардың ақуыздың денатурациясының алдын алудағы рөлі.
Шаперондар (III) суббірліктердің массасына қарай жіктеледі. Жоғары молекулалық шаперондардың массасы 60-тан 110 кД-ға дейін. Олардың ішінде ең көп зерттелген үш сынып: Ш-60, Ш-70 және Ш-90. Әрбір класқа бір-бірімен байланысты ақуыздар тобы кіреді. Осылайша, Sh-70 құрамында молекулалық салмағы 66-дан 78 кД-ға дейінгі ақуыздар бар. Төмен молекулалық салмақты шаперондардың молекулалық салмағы 40-15 кД болады.
Бақылаушылардың арасында бар құрылтайшыжоғары базальды синтезі дене жасушаларына стресстік әсерге байланысты емес ақуыздар және индукциялық,оның синтезі қалыпты жағдайда әлсіз, бірақ стресстік әсерде күрт артады. Индукцияланатын шаперондарды «жылу соққысының белоктары» деп те атайды, өйткені олар алғаш рет жоғары температураға ұшыраған жасушаларда табылған. Жасушаларда белоктардың жоғары концентрациясына байланысты жартылай денатуратталған белоктардың өздігінен регенерациялануы қиын. Sh-70 басталған денатурация процесін болдырмайды және белоктардың табиғи конформациясын қалпына келтіруге көмектеседі. Молекулярлық шаперондар-70- жасушаның барлық бөліктерінде: цитоплазмада, ядрода, эндоплазмалық ретикулумда, митохондрияда кездесетін жоғары дәрежеде сақталған белоктар класы. Sh-70 жалғыз полипептидтік тізбегінің карбоксил соңында ұзындықтағы пептидтермен әрекеттесе алатын ойық болып табылатын аймақ бар.
гидрофобты радикалдармен байытылған 7-ден 9-ға дейін аминқышқылдарының қалдықтары. Глобулярлы ақуыздардағы мұндай орындар шамамен әрбір 16 аминқышқылында кездеседі. Sh-70 ақуыздарды термиялық инактивациядан қорғауға және ішінара денатуратталған ақуыздардың конформациясы мен белсенділігін қалпына келтіруге қабілетті.
2. Шаперондардың белок қатпарлануындағы рөлі.Рибосомадағы белоктардың синтезі кезінде полипептидтің N-соңғы аймағы С-соңғы аймағынан бұрын синтезделеді. Нәтижелі конформацияны қалыптастыру үшін ақуыздың аминқышқылдарының толық тізбегі қажет. Ақуыз синтезі процесінде шаперондар-70 белсенді орталығының құрылымына байланысты синтез аяқталғанға дейін гидрофобты аминқышқылдарының радикалдарымен байытылған агрегацияға бейім полипептидтік учаскелерді жабуға қабілетті (1.27, А-сурет).
 Күріш. 1.27. Шаперондардың белок қатпарлануына қатысуы
Күріш. 1.27. Шаперондардың белок қатпарлануына қатысуы
А - синтезделген полипептидтің учаскелері арасындағы гидрофобты әрекеттесулердің алдын алуға шаперондар-70 қатысуы; В – шаперон кешенінде нативті белок конформациясының түзілуі
Домендік құрылым сияқты күрделі конформациясы бар көптеген жоғары молекулалық ақуыздар W-60 түзетін арнайы кеңістікте қатпарланады. Ш-60 14 бөлімшеден тұратын олигомерлі кешен қызметін атқарады. Олар екі қуыс сақина құрайды, олардың әрқайсысы жеті бөлімшеден тұрады, бұл сақиналар бір-бірімен байланысқан. III-60 әрбір суббірлігі үш доменнен тұрады: апикальды (апикальды), сақина қуысына қарайтын гидрофобты радикалдармен байытылған, аралық және экваторлық (1.28-сурет).
 Күріш. 1.28. Шаперонин кешенінің құрылымы 14 Ш-60
Күріш. 1.28. Шаперонин кешенінің құрылымы 14 Ш-60
A - бүйірлік көрініс; B - жоғарғы көрініс
Бүктелмеген молекулаларға тән беткі элементтері бар синтезделген белоктар, атап айтқанда, гидрофобты радикалдар шаперон сақиналарының қуысына енеді. Бұл қуыстардың ерекше ортасында мүмкін болатын конформацияларды санау жалғыз, энергетикалық жағынан ең қолайлысы табылғанға дейін жүреді (1.27, В-сурет). Конформациялардың түзілуі және ақуыздың бөлінуі экваторлық аймақта АТФ гидролизімен жүреді. Әдетте, шаперонға тәуелді мұндай бүктеу энергияның айтарлықтай мөлшерін талап етеді.
Шаперондар белоктардың үш өлшемді құрылымын қалыптастыруға және ішінара денатуратталған белоктарды ренатациялауға қатысудан басқа, олигомерлі белоктарды жинақтау, денатуратталған ақуыздарды лизосомаларға тану және тасымалдау, белоктарды тасымалдау сияқты іргелі процестерге де қажет. мембраналар арқылы және ақуыз кешендерінің қызметін реттеуге қатысады.
ТАҚЫРЫП 1.6. ПРОТЕИНДІҢ ТҮРЛІ. ИММУНОГЛОБУЛИНДЕР МЫСАЛЫНДАҒЫ БЕЛЕКТІЛІК ОТБАСЫ
1. Белоктар жеке жасушалардың және бүкіл көп жасушалы организмнің өмірінде шешуші рөл атқарады және олардың функциялары таңқаларлық әртүрлі. Бұл белоктардың біріншілік құрылымы мен конформациясының ерекшеліктерімен, белсенді орталықтың ерекше құрылымымен және нақты лигандтарды байланыстыру қабілетімен анықталады.
Пептидтік тізбектердің барлық мүмкін нұсқаларының өте аз ғана бөлігі тұрақты кеңістіктік құрылымды қабылдай алады; көпшілік
олардың шамамен бірдей Гиббс энергиясы бар, бірақ әртүрлі қасиеттері бар көптеген конформацияларды қабылдай алады. Белгілі белоктардың көпшілігінің бастапқы құрылымы, таңдалған биологиялық эволюция, осы ақуыздың жұмыс істеу ерекшеліктерін анықтайтын конформациялардың бірінің ерекше тұрақтылығын қамтамасыз етеді.
2. Белок тұқымдастары.Бір биологиялық түрдің ішінде аминқышқылдарының қалдықтарын алмастыру өзара байланысты функцияларды орындайтын және аминқышқылдарының гомологтық тізбегі бар әртүрлі ақуыздардың пайда болуына әкелуі мүмкін. Мұндай туыстас белоктардың таңғаларлық ұқсас конформациялары бар: α-спиральдардың және/немесе β-құрылымдардың саны мен орналасуы, және полипептидтік тізбектердің бұрылыстары мен қатпарларының көпшілігі ұқсас немесе бірдей. Полипептидтік тізбектің гомологты аймақтары, ұқсас конформациясы және соған байланысты функциялары бар белоктар белоктар тұқымдастарына бөлінеді. Белок тұқымдастарының мысалдары: сериндік протеиназалар, иммуноглобулиндер семьясы, миоглобиндер семьясы.
Сериндік протеиназалар- протеолитикалық ферменттер қызметін атқаратын белоктар семьясы. Оларға ас қорыту ферменттері – химотрипсин, трипсин, эластаза және қанның ұюының көптеген факторлары жатады. Бұл белоктардың 40% бірдей аминқышқылдары және конформациясы өте ұқсас (1.29-сурет).

Күріш. 1.29. Эластазаның (А) және химотрипсиннің (В) кеңістіктік құрылымдары
Кейбір аминқышқылдарының алмастырулары осы белоктардың субстрат ерекшелігінің өзгеруіне және отбасында функционалды әртүрліліктің пайда болуына әкелді.
3. Иммуноглобулиндер тұқымдасы.Иммундық жүйенің жұмысында үлкен рөлбелоктардың үш тобын қамтитын иммуноглобулиндердің суперфамилиясының протеиндерін ойнайды:
антиденелер (иммуноглобулиндер);
Т-лимфоциттердің рецепторлары;
Негізгі гистосәйкестік кешенінің белоктары – MHC 1 және 2 кластар (Негізгі гистосәйкестік кешені).
Бұл белоктардың барлығы домендік құрылымға ие, гомологтық иммундық тәрізді домендерден тұрады және ұқсас қызметтерді орындайды: олар қанда, лимфада немесе жасушааралық сұйықтықта (антиденелер) еріген бөтен құрылымдармен әрекеттеседі немесе жасушалардың бетінде (меншікті немесе шетелдік).
4. Антиденелер- деп аталатын бөгде құрылымды жұту жауап В-лимфоциттер өндірілген арнайы белоктар антиген.
Антиденелердің құрылысының ерекшеліктері
Қарапайым антидене молекулалары төрт полипептидтік тізбектен тұрады: екі бірдей жеңіл тізбек – L, құрамында шамамен 220 амин қышқылдары және екі бірдей ауыр тізбек – H, 440-700 аминқышқылдарынан тұрады. Антидене молекуласындағы төрт тізбектің барлығы да көптеген ковалентті емес байланыстар және төрт дисульфидтік байланыстар арқылы байланысқан (1.30-сурет).
Антиденелердің жеңіл тізбектері екі доменнен тұрады: айнымалы (VL), полипептидтік тізбектің N-соңғы аймағында орналасқан және тұрақты (CL), С-соңында орналасқан. Ауыр тізбектер әдетте төрт доменге ие: N-терминуста бір айнымалы (VH) және үш тұрақты (CH1, CH2, CH3) (1.30-суретті қараңыз). Әрбір иммуноглобулин доменінде екі цистеин қалдығы дисульфидтік байланыспен байланысқан β-бүрінді қондырма бар.
Екі тұрақты CH1 және CH2 домендерінің арасында екінші реттік құрылымның түзілуіне және осы сегменттегі көршілес Н-тізбектерінің өзара әрекеттесуіне жол бермейтін көптеген пролин қалдықтары бар аймақ бар. Бұл топса аймағы антидене молекуласына икемділік береді. Ауыр және жеңіл тізбектердің айнымалы домендерінің арасында екі бірдей антигенді байланыстыру орындары (антигендерді байланыстыратын белсенді учаскелер) орналасқан, сондықтан мұндай антиденелер жиі деп аталады. биваленттер.Антигеннің антиденемен байланысуы екі тізбектің айнымалы аймақтарының барлық аминқышқылдарының тізбегін қамтымайды, бірақ әрбір тізбектің гипервариативті аймақтарында орналасқан 20-30 аминқышқылдары ғана болады. Дәл осы аймақтар антиденелердің әрбір түрінің сәйкес комплементарлы антигенмен әрекеттесуінің бірегей қабілетін анықтайды.
Антиденелер - ағзаның бөтен организмдердің енуіне қарсы қорғаныс желілерінің бірі. Олардың жұмыс істеуін екі кезеңге бөлуге болады: бірінші кезең - антидене құрылымында антигенді байланыстыру орындарының болуына байланысты мүмкін болатын бөгде организмдердің бетіндегі антигенді тану және байланыстыру; екінші кезең - антигенді инактивациялау және жою процесінің басталуы. Екінші кезеңнің ерекшелігі антиденелер класына байланысты. Тұрақты домендердің құрылымы бойынша бір-бірінен ерекшеленетін ауыр тізбектердің бес класы бар: α, δ, ε, γ және μ, соған сәйкес иммуноглобулиндердің бес класы бөлінеді: A, D, E, G және M.
Ауыр тізбектердің құрылымдық ерекшеліктері ауыр тізбектердің топса аймақтары мен С-терминалды аймақтарына әрбір класқа тән конформациялық сипаттама береді. Антиген антиденемен байланысқаннан кейін, тұрақты домендердегі конформациялық өзгерістер антигенді жою жолын анықтайды.
 Күріш. 1. 30. IgG домендік құрылымы
Күріш. 1. 30. IgG домендік құрылымы
Иммуноглобулиндер М
Иммуноглобулиндер М екі нысаны бар.
Мономерлі форма- дамып келе жатқан В-лимфоцит өндіретін антиденелердің 1-ші класы. Кейіннен көптеген В жасушалары антиденелердің басқа кластарын өндіруге ауысады, бірақ антигенді байланыстыру орны бірдей. IgM мембранаға еніп, антигенді тану рецепторы қызметін атқарады. IgM жасуша мембранасына қосылуы аймақтың құйрық бөлігінде 25 гидрофобты амин қышқылы қалдықтарының болуына байланысты мүмкін.
IgM секреторлық формасыбір-бірімен дисульфидтік байланыспен байланысқан бес мономерлі суббірліктер және қосымша полипептидті J-тізбектен тұрады (1.31-сурет). Бұл түрдегі ауыр тізбекті мономерлерде гидрофобты құйрық болмайды. Пентамердің 10 антигенді байланыстыратын орны бар, сондықтан организмге бірінші рет енген антигенді тану және жоюда тиімді. IgM секреторлық түрі біріншілік иммундық жауап кезінде қанға бөлінетін антиденелердің негізгі класы болып табылады. IgM-нің антигенмен байланысуы IgM конформациясын өзгертеді және оның комплемент жүйесінің бірінші белоктық компонентімен байланысуын (комплемент жүйесі – антигеннің бұзылуына қатысатын белоктардың жиынтығы) және осы жүйенің активтенуін индукциялайды. Егер антиген микроорганизмнің бетінде орналасса, комплемент жүйесі тұтастығын бұзады. жасуша мембранасыжәне бактерия жасушасының өлуі.
Иммуноглобулиндер Г
Сандық тұрғыдан алғанда, иммуноглобулиндердің бұл класы қанда басым (барлық Ig-нің 75%). IgG – мономерлер, екіншілік иммундық жауап кезінде қанға бөлінетін антиденелердің негізгі класы. IgG микроорганизмдердің беткі антигендерімен әрекеттескеннен кейін антиген-антидене кешені комплемент жүйесінің ақуыздарын байланыстыруға және белсендіруге қабілетті немесе макрофагтар мен нейтрофилдердегі арнайы рецепторлармен әрекеттесе алады. фагоциттермен әрекеттесу
 Күріш. 1.31. IgM секреторлық формасының құрылымы
Күріш. 1.31. IgM секреторлық формасының құрылымы
антиген-антидене кешендерінің сіңуіне және олардың жасушалардың фагосомаларындағы жойылуына. IgG - бұл плацентарлы тосқауылдан өтіп, ұрықты жатырдағы инфекциялардан қорғай алатын жалғыз антиденелер класы.
Иммуноглобулиндер А
Секрецияда болатын антиденелердің негізгі класы (сүт, сілекей, тыныс алу және ішек секрециялары). IgA негізінен димерлі түрде бөлінеді, мұнда мономерлер бір-бірімен қосымша J-тізбек арқылы байланысады (1.32-сурет).
IgA комплемент жүйесімен және фагоцитарлық жасушалармен әрекеттеспейді, бірақ микроорганизмдермен байланысу арқылы антиденелер олардың эпителий жасушаларына қосылып, организмге енуіне жол бермейді.
Иммуноглобулиндер Е
Е иммуноглобулиндері ауыр ε-тізбектері бар мономерлермен, сондай-ақ М иммуноглобулиндерінің μ-тізбектерімен, бір айнымалы және төрт тұрақты доменмен ұсынылған. IgE секрециядан кейін өздерімен байланысады
 Күріш. 1.32. IgA құрылымы
Күріш. 1.32. IgA құрылымы
Мастикалық жасушалар мен базофилдердің бетінде сәйкес рецепторлары бар С-терминалды аймақтар. Нәтижесінде олар осы жасушалардың бетіндегі антигендердің рецепторларына айналады (1.33-сурет).
 Күріш. 1.33. IgE-нің антигенмен мастикалық жасуша бетіндегі әрекеттесуі
Күріш. 1.33. IgE-нің антигенмен мастикалық жасуша бетіндегі әрекеттесуі
Антиген сәйкес антигенді байланыстыратын IgE учаскелеріне бекітілгеннен кейін жасушалар қабыну реакциясының дамуына және аллергиялық реакциялардың көрінісіне жауап беретін биологиялық белсенді заттардың (гистамин, серотонин) бөлінуіне сигнал алады. демікпе, есекжем, шөп безгегі.
Иммуноглобулиндер D
D иммуноглобулиндері сарысуда өте аз мөлшерде кездеседі, олар мономерлер. Ауыр δ тізбектерде бір айнымалы және үш тұрақты домен болады. IgD В-лимфоциттердің рецепторлары ретінде әрекет етеді, басқа функциялары әлі белгісіз. Арнайы антигендердің В-лимфоциттердің (IgD) бетіндегі рецепторлармен әрекеттесуі бұл сигналдардың жасушаға берілуіне және лимфоциттердің осы клонының көбеюін қамтамасыз ететін механизмдердің белсендірілуіне әкеледі.
ТАҚЫРЫП 1.7. БЕЛГІЛЕРДІҢ ФИЗИКАЛЫҚ-ХИМИЯЛЫҚ ҚАСИЕТТЕРІ ЖӘНЕ ОЛАРДЫ БӨЛУ ӘДІСТЕРІ.
1. Жеке белоктар физикалық-химиялық қасиеттері бойынша ерекшеленеді:
Молекулалардың пішіні;
Молекулярлық салмақ;
Жалпы заряд, оның мәні аминқышқылдарының анионды және катиондық топтарының қатынасына байланысты;
Молекулалар бетіндегі полярлы және полярсыз аминқышқылдарының радикалдарының қатынасы;
Әртүрлі денатурациялаушы агенттерге төзімділік дәрежелері.
2. Белоктардың ерігіштігі тәуелдіжоғарыда аталған белоктардың қасиеттері туралы, сондай-ақ белок еритін ортаның құрамы туралы (рН мәндері, тұз құрамы, температура, ақуызбен әрекеттесе алатын басқа органикалық заттардың болуы). Белок молекулаларының зарядының шамасы олардың ерігіштігіне әсер ететін факторлардың бірі болып табылады. Заряд изоэлектрлік нүктеде жоғалған кезде белоктар оңайырақ біріктіріліп, тұнбаға түседі. Бұл әсіресе бетінде гидрофобты аминқышқылдарының радикалдары бар денатуратталған ақуыздарға қатысты.
Белок молекуласының бетінде оң және теріс зарядталған аминқышқылдарының радикалдары болады. Бұл топтардың саны, демек, белоктардың жалпы заряды ортаның рН-ына байланысты, яғни. Н + - және ОН - топтарының концентрациясының қатынасы. Қышқыл ортада H+ концентрациясының жоғарылауы карбоксил топтарының -COO - + H+ > -COOH диссоциациясының басылуына және белоктардың теріс зарядының төмендеуіне әкеледі. Сілтілік ортада амин топтарының -NH 3 + + OH - - NH 2 + H 2 O диссоциациясы кезінде түзілетін артық OH - протондардың судың түзілуімен байланысуы белоктардың оң зарядының төмендеуіне әкеледі. Ақуыздың таза заряды нөлге тең болатын рН мәні деп аталады изоэлектрлік нүкте (IEP). IET-те оң және теріс зарядталған топтардың саны бірдей, яғни. белок изоэлектрлік күйде болады.
3. Жеке белоктардың бөлінуі.Организмнің құрылымы мен қызмет ету ерекшеліктері онда синтезделген белоктардың жиынтығына байланысты. Белоктардың құрылысы мен қасиеттерін зерттеу оларды жасушадан бөліп алып, басқа белоктар мен органикалық молекулалардан тазартпай мүмкін емес. Жеке белоктарды бөліп алу және тазарту кезеңдері:
жасушаның жойылуызерттелетін тіннің және гомогенатты алу.
Гомогенатты фракцияларға бөлуцентрифугалау, қажетті ақуызды қамтитын ядролық, митохондриялық, цитозолдық немесе басқа фракцияны алу.
Селективті жылулық денатурация- белок ерітіндісін қысқа мерзімді қыздыру, онда денатуратталған ақуыз қоспаларының бір бөлігін жоюға болады (ақуыз салыстырмалы түрде термиялық тұрақты болған жағдайда).
Тұздану.Әртүрлі ақуыздар ерітіндідегі тұздың әртүрлі концентрациясында тұнбаға түседі. Тұз концентрациясын бірте-бірте арттыра отырып, олардың біреуінде бөлінетін белок басым болатын бірқатар жеке фракцияларды алуға болады. Белоктардың ең көп қолданылатын фракциясы аммоний сульфаты болып табылады. Төмен ерігіштігі бар белоктар тұздың төмен концентрациясында тұнбаға түседі.
Гельді сүзу- ісінген Sephadex түйіршіктері (кеуектері бар үш өлшемді декстран полисахаридті тізбектер) арқылы молекулаларды елеуіш әдісі. Белоктардың Sephadex толтырылған колонка арқылы өту жылдамдығы олардың молекулалық салмағына байланысты болады: ақуыз молекулаларының массасы неғұрлым аз болса, олар түйіршіктерге оңай еніп, оларда ұзағырақ қалады, массасы неғұрлым үлкен болса, олар соғұрлым тезірек бөлінеді. баған.
Ультрацентрифугалау- центрифугалық түтіктегі ақуыздардың ультрацентрифуганың роторына орналастырылуынан тұратын әдіс. Ротор айналу кезінде ақуыздың шөгу жылдамдығы оларға пропорционал болады молекулалық салмақ: ауыр белоктардың фракциялары түтік түбіне жақын орналасады, жеңілдері бетіне жақын орналасады.
электрофорез- электр өрісіндегі белоктардың қозғалыс жылдамдығының айырмашылығына негізделген әдіс. Бұл мән белоктардың зарядына пропорционал. Ақуыз электрофорезі қағазда (бұл жағдайда белоктардың қозғалу жылдамдығы тек олардың зарядына пропорционалды) немесе белгілі бір кеуек өлшемі бар полиакриламидті гельде (белоктардың қозғалыс жылдамдығы олардың заряды мен молекулалық массасына пропорционал) жүргізіледі. ).
Ион алмасу хроматографиясы- белоктардың иондалған топтарын ион алмастырғыш шайырлардың (ерімейтін полимерлі материалдар) қарама-қарсы зарядталған топтарымен байланыстыруға негізделген фракциялау әдісі. Ақуыздың шайырмен байланысу күші ақуыздың зарядына пропорционал. Ион алмастырғыш полимерге адсорбцияланған белоктарды NaCl ерітінділерінің концентрациясының жоғарылауымен жууға болады; ақуыз заряды неғұрлым төмен болса, шайырдың иондық топтарымен байланысқан ақуызды жуу үшін NaCl концентрациясының төмен болуы қажет болады.
Сәйкестік хроматографиясы- жеке белоктарды бөліп алудың ең спецификалық әдісі.Белоктың лигандасы инертті полимерге ковалентті байланысады. Белок ерітіндісін полимері бар колонка арқылы өткізгенде, белоктың лигандпен комплементарлы байланысуына байланысты колоннаға тек осы лигандқа тән ақуыз ғана адсорбцияланады.
Диализ- оқшауланған ақуыз ерітіндісінен төмен молекулалық қосылыстарды жою үшін қолданылатын әдіс. Әдіс белоктардың төмен молекулалы заттарға қарағанда жартылай өткізгіш мембранадан өте алмауына негізделген. Ол ақуыздарды төмен молекулалық қоспалардан, мысалы, тұздан кейін тұздардан тазарту үшін қолданылады.
СЫНЫПТАН ТЫС ЖҰМЫСТАРҒА ТАПСЫРМАЛАР
1. Кестені толтыру. 1.4.
1.4-кесте. Салыстырмалы талдаутуыстас белоктардың – миоглобин мен гемоглобиннің құрылымдары мен қызметтері
а) белсенді орталықтың құрылымын есте сақтаңыз Mb және Hb. Аминқышқылдарының гидрофобты радикалдары осы белоктардың белсенді орталықтарының түзілуінде қандай рөл атқарады? Mb және Hb белсенді орталығының құрылымын және оған лигандтардың қосылу механизмдерін сипаттаңыз. Оның F 8 және Оның E 7 қалдықтары Mv және Hv белсенді учаскесінің қызметінде қандай рөл атқарады?
б) мономерлі миоглобинмен салыстырғанда бір-бірімен тығыз байланысты олигомерлі ақуыз гемоглобиннің қандай жаңа қасиеттері бар? Гемоглобин молекуласындағы протомерлердің конформациясындағы кооперативтік өзгерістердің рөлін, СО 2 мен протон концентрациясының гемоглобиннің оттегіге жақындығына әсерін және Hb функциясының аллостериялық реттелуіндегі 2,3-ВПГ рөлін түсіндіріңіз.
2. Молекулярлық шаперондардың құрылымы мен қызметі арасындағы байланысқа назар аудара отырып, олардың сипаттамаларын сипаттаңыз.
3. Қандай белоктар тұқымдастарға топтастырылған? Иммуноглобулиндер тұқымдасына мысал келтіре отырып, осы тұқымдас белоктардың ұқсас құрылымдық ерекшеліктерін және оларға байланысты қызметтерін анықтаңыз.
4. Көбінесе биохимиялық және медициналық мақсаттар үшін тазартылған жеке ақуыздар қажет. Белоктардың қандай физика-химиялық қасиеттеріне негізделген оларды бөлу және тазарту әдістерін түсіндіріңіз.
ӨЗІН-ӨЗІ БАҚЫЛАУ Тапсырмалары
1. Дұрыс жауаптарды таңдаңыз.
Гемоглобиннің функциялары:
A. O 2 өкпеден ұлпаларға тасымалданады B. H + ұлпалардан өкпеге тасымалданады
B. Тұрақты қан рН деңгейін ұстап тұру D. СО2 өкпеден ұлпаларға тасымалдануы
D. СО 2-нің ұлпалардан өкпеге тасымалдануы
2. Дұрыс жауаптарды таңдаңыз. лигандα -Hb протомері:А. Хеме
B. Оттегі
B. CO D. 2,3-BPG
D. β-протомер
3. Дұрыс жауаптарды таңдаңыз.
Гемоглобиннің миоглобиннен айырмашылығы:
A. Төрттік құрылымы бар
B. Қосалқы құрылым тек α-спиральдармен берілген
B. Күрделі белоктарға жатады
D. Аллостериялық лигандпен әрекеттеседі D. Геммен ковалентті байланысқан
4. Дұрыс жауаптарды таңдаңыз.
Hb-ның O 2-ге жақындығы төмендейді:
A. Бір O 2 молекуласы қосылғанда B. Бір O 2 молекуласы жойылғанда
B. 2,3-BPG-мен әрекеттесу кезінде
D. H+D протомерлеріне қосылғанда. 2,3-БПГ концентрациясы төмендегенде
5. Сәйкестік орнату.
Hb типтері үшін ол тән:
A. Дезокси түрінде фибриллярлы агрегаттар түзеді B. Құрамында екі α- және екі δ-тізбектері бар.
B. Ересек эритроциттерде Hb басым түрі D. Белсенді орталығында Fe+3 бар гем бар.
D. Құрамында екі α- және екі γ-тізбектері бар 1. HvA 2.
6. Сәйкестік орнату.
Лигандалар Hb:
A. Аллостериялық орталықта Hb-мен байланысады
B. Белсенді Hb сайтына өте жоғары жақындығы бар
B. Қосылу, Hb-ның O 2-ге жақындығын арттырады D. Fe + 2-ні Fe + 3-ке тотықтырады.
D. Пішіндер коваленттік байланыс hysF8 көмегімен
7. Дұрыс жауаптарды таңдаңыз.
Каперондар:
A. Жасушаның барлық бөліктерінде болатын белоктар
B. Стресстік әсерлер кезінде синтез күшейеді
B. Денатуратталған белоктардың гидролизіне қатысады
D. Белоктардың нативті конформациясын сақтауға қатысу
D. Белок конформациясы түзілетін органоидтарды жасаңыз
8. Сәйкестік. Иммуноглобулиндер:
A. Секреторлық формасы бесжылдық
B. Плацентарлы тосқауылдан өтетін Ig класы
B. Ig - діңгек жасушаларының рецепторы
D. Эпителий жасушаларының секрецияларында болатын Ig негізгі класы. D. В-лимфоциттік рецептор, оның активтенуі жасушаның көбеюін қамтамасыз етеді
9. Дұрыс жауаптарды таңдаңыз.
Иммуноглобулиндер Е:
A. Макрофагтар түзеді B. Ауыр ε-тізбектері бар.
B. Т-лимфоциттердің жарғақшасына енген
D. Мастикалық жасушалар мен базофилдердегі антигендер үшін мембраналық рецепторлар ретінде әрекет етеді
D. Аллергиялық реакциялардың көрінісіне жауапты
10. Дұрыс жауаптарды таңдаңыз.
Белоктарды бөлу әдісі олардың молекулалық салмағының айырмашылығына негізделген:
A. Гельді сүзу
B. Ультрацентрифугалау
B. Полиакриламидті гельдік электрофорез D. Ион алмасу хроматографиясы
D. Афинитті хроматография
11. Дұрыс жауапты таңда.
Ақуыздарды бөлу әдісі олардың судағы ерігіштігінің айырмашылығына негізделген:
A. Гельді сүзу B. Тұздану
B. Ион алмасу хроматографиясы D. Афинитті хроматография
E. Полиакриламидті гельді электрофорез
«ӨЗІН-ӨЗІ БАҚЫЛАУ ТАПСЫРМАЛАРЫНА» ЖАУАП БЕРУ СТАНДАРТтары
1. A, B, C, D
2. A, B, C, D
5. 1-В, 2-А, 3-Д
6. 1-С, 2-Б, 3-А
7. A, B, D, D
8. 1-G; 2-В, 3-С
НЕГІЗГІ ТЕРМИНДЕР МЕН ТҮСІНІКТЕР
1. Олигомерлі белоктар, протомер, белоктардың төрттік құрылымы
2. Протомер конформациясындағы кооперативтік өзгерістер
3. Бор эффектісі
4. Белок функцияларының аллостериялық реттелуі, аллостериялық орталық және аллостериялық эффектор
5. Молекулярлық шаперондар, жылу соққысының белоктары
6. Белок тұқымдастары (сериндік протеазалар, иммуноглобулиндер)
7. IgM-, G-, E-, A- құрылымның функциямен байланысы
8. Белоктардың толық заряды, белоктардың изоэлектрлік нүктесі
9. Электрофорез
10. Тұзды шығару
11. Гельді фильтрация
12. Ион алмасу хроматографиясы
13. Ультрацентрифугалау
14. Афинитті хроматография
15. Плазма ақуызының электрофорезі
АУДИЦИЯ ЖҰМЫСЫНА АРНАЛҒАН ТАПСЫРМАЛАР
1. Гемоглобин (Нв) мен миоглобиннің (Мб) оттегімен қанығу дәрежесінің оның ұлпалардағы парциалды қысымына тәуелділігін салыстырыңыз.
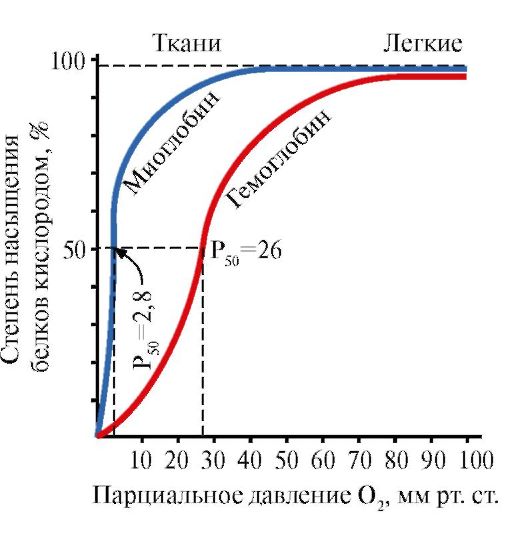 Күріш. 1.34. МВ-ның қанығуға тәуелділігі жәнеHbоның парциалды қысымынан оттегі
Күріш. 1.34. МВ-ның қанығуға тәуелділігі жәнеHbоның парциалды қысымынан оттегі
Ақуыздың оттегімен қанығу қисықтарының пішіні әртүрлі екенін ескеріңіз: миоглобин үшін - гипербола, гемоглобин үшін - сигма тәрізді пішін.
1. Mb және Hb O 2-ге 50% қаныққан оттегінің парциалды қысымының мәндерін салыстырыңыз. Осы белоктардың қайсысының O 2-ге жақындығы жоғары?
2. МБ қандай құрылымдық ерекшеліктері оның O 2-ге жоғары жақындығын анықтайды?
3. Hb-ның қандай құрылымдық ерекшеліктері тыныштықтағы тіндердің капиллярларында (О 2 салыстырмалы жоғары парциалды қысымда) О 2-ні босатуға және жұмыс істейтін бұлшықеттерде бұл қайтарымды күрт арттыруға мүмкіндік береді? Бұл әсерді олигомерлі белоктардың қандай қасиеті қамтамасыз етеді?
4. Тыныштықтағы және жұмыс істейтін бұлшықетке оттегімен қаныққан гемоглобинді қандай мөлшерде О 2 (%-бен) беретінін есептеңіз?
5. ақуыз құрылымы мен оның қызметі арасындағы байланыс туралы қорытынды жасау.
2. Капиллярлардағы гемоглобин бөлетін оттегінің мөлшері тіндердегі катаболизм процестерінің қарқындылығына байланысты (Бор эффектісі). Тіндердің метаболизміндегі өзгерістер Hb-ның O 2-ге жақындығын қалай реттейді? CO 2 және H+ Hb-ның O 2-ге жақындығына әсері
 1. Бор эффектісін сипаттаңыз.
1. Бор эффектісін сипаттаңыз.
2. диаграммада көрсетілген процесс қай бағытта жүреді?
а) өкпе капиллярларында;
б) ұлпа капиллярларында?
3. Бор эффектінің физиологиялық маңызы қандай?
4. Неліктен гемнен қашық орналасқан жерлерде Hb-ның Н+-мен әрекеттесуі ақуыздың O 2-ге жақындығын өзгертеді?
3. Hb-ның O 2-ге жақындығы оның лигандының, 2,3-бифосфоглицераттың концентрациясына байланысты, ол Hb-ның O 2-ге жақындығының аллостериялық реттеушісі болып табылады. Неліктен белсенді аймақтан қашықтағы тораптағы лигандтардың өзара әрекеттесуі ақуыз қызметіне әсер етеді? 2,3-BPG Hb-ның O 2-ге жақындығын қалай реттейді? Мәселені шешу үшін келесі сұрақтарға жауап беріңіз:
1. 2,3-бифосфоглицерат (2,3-БПГ) қайдан және неден синтезделеді? Оның формуласын жаз, осы молекуланың зарядын көрсет.
2. БПГ гемоглобиннің қандай формасымен (окси немесе дезокси) әрекеттеседі және неліктен? Hb молекуласының қай аймағында әрекеттесу жүреді?
3. сызбада көрсетілген процесс қай бағытта жүреді?
а) ұлпа капиллярларында;
б) өкпе капиллярларында?
4. қай жерде көбірек болуы керек жоғары концентрациякешен
Nv-2,3-BFG:
а) тыныштықтағы бұлшықет капиллярларында;
б) жұмыс істейтін бұлшықеттердің капиллярларында (эритроциттерде BPG бірдей концентрациясын есептегенде)?
5. Эритроциттердегі БПГ концентрациясы жоғарыласа, адам биіктік жағдайына бейімделген кезде Hb оттегіге жақындығы қалай өзгереді? Бұл құбылыстың физиологиялық маңызы қандай?
4. Консервіленген қанды сақтау кезінде 2,3-БПГ-ның бұзылуы Hb функцияларын бұзады. Егер эритроциттердегі 2,3-БПГ концентрациясы 8-ден 0,5 ммоль/л-ге дейін төмендеуі мүмкін болса, сақталған қандағы Hb-ның O 2-ге жақындығы қалай өзгереді. 2,3-БПГ концентрациясы үш күннен ерте емес қалпына келтірілсе, ауыр науқастарға мұндай қанды құюға болады ма? Қанға 2,3-БПГ қосу арқылы эритроциттердің қызметін қалпына келтіруге болады ма?
5. Ең қарапайым иммуноглобулин молекулаларының құрылымын еске түсіріңіз. Иммуноглобулиндер иммундық жүйеде қандай рөл атқарады? Неліктен Igs жиі бивалентті деп аталады? Igs құрылымы олардың қызметімен қалай байланысты? (Иммуноглобулиндер класының мысалын пайдаланып сипаттаңыз.)
Белоктардың физика-химиялық қасиеттері және оларды бөлу әдістері.
6. Ақуыздың таза заряды оның ерігіштігіне қалай әсер етеді?
а) рН 7 кезінде пептидтің жалпы зарядын анықтаңыз
Ala-Glu-Tre-Pro-Asp-Liz-Cis
б) рН >7, рН кезінде осы пептидтің заряды қалай өзгереді<7, рН <<7?
в) белоктың изоэлектрлік нүктесі (ИЭП) қандай және ол қандай ортада орналасады
Осы пептидтің IET?
г) қандай рН шамасында осы пептидтің ең аз ерігіштігі байқалады.
7. Неліктен қышқыл сүт, жаңа піскен сүтке қарағанда, қайнатқанда «ұйытады» (яғни, казеин сүт протеині тұнбаға түседі)? Жаңа сүттегі казеин молекулаларының теріс заряды бар.
8. Гельді фильтрация жеке белоктарды бөлу үшін қолданылады. Молекулярлық массалары сәйкесінше 160 000, 80 000 және 60 000-ға тең құрамында А, В, С белоктары бар қоспа гельді фильтрация арқылы талданды (1.35-сурет). Ісінген гель түйіршіктері молекулалық массасы 70000-нан аз белоктарды өткізеді.Бұл бөлу әдісінің негізінде қандай принцип жатыр? Графиктердің қайсысы бөлшектеу нәтижелерін дұрыс көрсетеді? А, В және С белоктарының бағаннан шығу ретін көрсетіңіз.
 Күріш. 1.35. Протеиндерді бөлу үшін гельді сүзу әдісін пайдалану
Күріш. 1.35. Протеиндерді бөлу үшін гельді сүзу әдісін пайдалану
9. Суретте. 1.36, А сау адамның қан сарысуындағы ақуыздардың қағаздағы электрофорез сызбасын көрсетеді. Бұл әдіс арқылы алынған ақуыз фракцияларының салыстырмалы мөлшері: альбуминдер 54-58%, α 1 -глобулиндер 6-7%, α 2 -глобулиндер 8-9%, β-глобулиндер 13%, γ-глобулиндер 11-12% .
 Күріш. 1.36 Сау адамның (А) және науқастың (В) қан плазмасының ақуыздарының қағаздағы электрофорезі
Күріш. 1.36 Сау адамның (А) және науқастың (В) қан плазмасының ақуыздарының қағаздағы электрофорезі
I – γ-глобулиндер; II – β-глобулиндер; III -α 2 - глобулин; IV-α 2 - глобулин; V – альбуминдер
Көптеген аурулар сарысу ақуыздарының құрамындағы сандық өзгерістермен (диспротеинемия) жүреді. Бұл өзгерістердің сипаты диагноз қою және аурудың ауырлығы мен сатысын бағалау кезінде ескеріледі.
Кестеде берілген мәліметтерді пайдалану. 1.5, суретте көрсетілген электрофоретикалық профильмен сипатталатын ауру туралы болжам жасаңыз. 1.36.
1.5-кесте. Патологияда қан сарысуындағы ақуыздардың концентрациясының өзгеруі
5. Реттеуші функция. Белоктар сигналдық заттардың функцияларын орындайды - кейбір гормондар, гистогормондар және нейротрансмиттерлер, кез келген құрылымның сигналдық заттарының рецепторлары болып табылады, жасушаның биохимиялық сигналдық тізбектерінде одан әрі сигнал беруді қамтамасыз етеді. Мысалы, өсу гормоны соматотропин, инсулин гормоны, Н- және М-холинергиялық рецепторлар.
6. Қозғалтқыштың қызметі. Белоктардың көмегімен жиырылу және басқа да биологиялық қозғалыс процестері жүзеге асырылады. Мысалы, тубулин, актин, миозин.
7. Қосалқы функция. Өсімдіктерде құнды қоректік заттар болып табылатын сақтаушы ақуыздар бар, жануарларда бұлшық ет ақуыздары төтенше жағдайда жұмылдырылатын қоректік заттар қызметін атқарады.
Белоктар құрылымдық ұйымның бірнеше деңгейлерінің болуымен сипатталады.
бастапқы құрылымАқуыз - бұл полипептидтік тізбектегі аминқышқылдарының қалдықтарының тізбегі. Пептидтік байланыс – бір амин қышқылының α-карбоксил тобы мен басқа амин қышқылының α-амин тобы арасындағы карбоксамидті байланыс.
аланилфенилаланилцистеилпролин
U n эпидтік байланысбірнеше ерекшеліктері бар:
а) ол резонанстық тұрақтандырылған, сондықтан іс жүзінде бір жазықтықта орналасқан – ол жазық; C-N байланысының айналасында айналу көп энергияны қажет етеді және қиын;
б) -CO-NH- байланысы ерекше сипатқа ие, ол жайдан аз, бірақ екі еседен көп, яғни кетоэнол таутомериясы бар:

в) пептидтік байланысқа қатысты орынбасарлар транс-позиция;
г) пептидтік магистраль әртүрлі табиғаттағы бүйірлік тізбектермен қоршалған, қоршаған еріткіш молекулаларымен әрекеттесе отырып, бос карбоксил және амин топтары иондалып, белок молекуласының катиондық және аниондық орталықтарын құрайды. Олардың қатынасына байланысты белок молекуласы жалпы оң немесе теріс заряд алады, сонымен қатар ақуыздың изоэлектрлік нүктесіне жеткенде ортаның сол немесе басқа рН мәнімен сипатталады. Радикалдар белок молекуласының ішінде тұз, эфир, дисульфидті көпірлер түзеді, сонымен қатар белоктарға тән реакциялар ауқымын анықтайды.
Қазіргі уақытта 100 және одан да көп аминқышқылдарының қалдықтарынан тұратын полимерлерді белоктар, 50-100 аминқышқылдарының қалдықтарынан тұратын полимерлерді полипептидтер, ал 50-ден аз аминқышқылдарының қалдықтарынан тұратын полимерлерді төмен молекулалық пептидтер ретінде қарастыру туралы келісім жасалды.
Кейбір төмен молекулалық салмақпептидтер тәуелсіз биологиялық рөл атқарады. Осы пептидтердің кейбірінің мысалдары:
Глутатион - γ-glu-cis-gli - бірең кең таралған жасушаішілік пептидтердің бірі, ол жасушалардағы тотығу-тотықсыздану процестеріне және аминқышқылдарының биологиялық мембраналар арқылы тасымалдануына қатысады.
Карнозин - β-ала-гис - пептид,жануарлардың бұлшықеттерінде болады, липидтердің асқын тотығу өнімдерін жояды, бұлшықеттердегі көмірсулардың ыдырауын тездетеді және фосфат түрінде бұлшықеттердегі энергия алмасуына қатысады.
Вазопрессин – гипофиздің артқы бөлігінің гормоны, ағзадағы су алмасуын реттеуге қатысады:

Фаллоидин- улы шыбын агарикалық полипептид, шамалы концентрацияда жасушалардан ферменттер мен калий иондарының бөлінуіне байланысты организмнің өліміне әкеледі:

Грамицидин - антибиотик, көптеген грам-позитивті бактерияларға әсер ете отырып, төмен молекулалық қосылыстар үшін биологиялық мембраналардың өткізгіштігін өзгертеді және жасуша өлімін тудырады:

кездесті-энкефалин - тир-гли-гли-фен-мет - нейрондарда синтезделетін және ауырсынуды басатын пептид.
Белоктың екіншілік құрылымы- бұл пептидтік магистральдың функционалдық топтары арасындағы өзара әрекеттесу нәтижесінде пайда болатын кеңістіктік құрылым.
Пептидтік тізбектің құрамына кіредіпептидтік байланыстың көптеген CO және NH топтары, олардың әрқайсысы сутегі байланыстарының түзілуіне қатысуға қабілетті. Бұған мүмкіндік беретін құрылымдардың екі негізгі түрі бар: тізбек катушкалары телефон сымын ұнататын α-спираль және бір немесе бірнеше тізбектердің ұзартылған бөліктері қатар орналасқан β-бүйірлі құрылым. Бұл екі құрылым да өте тұрақты.
α-Helix сипатталадыбұралған полипептидтік тізбектің өте тығыз оралуы, оң жақ спиральдың әрбір айналымы үшін 3,6 амин қышқылы қалдықтары болады, олардың радикалдары әрқашан сыртқа және сәл артқа, яғни полипептидтік тізбектің басына бағытталған.
α-спиральдың негізгі сипаттамалары:
1) α-спираль пептидтік топтың азотындағы сутегі атомы мен қалдықтың карбонил оттегі арасындағы сутектік байланыстар арқылы тізбек бойынша берілгеннен төрт позицияда тұрақталады;
2) барлық пептидтік топтар α-спиральдың максималды тұрақтылығын қамтамасыз ететін сутектік байланыстың түзілуіне қатысады;
3) пептидтік топтардың барлық азот және оттегі атомдары сутектік байланыстардың түзілуіне қатысады, бұл α-спиральді аймақтардың гидрофильділігін айтарлықтай төмендетеді және олардың гидрофобтылығын арттырады;
4) α-спираль өздігінен түзіледі және бос энергияның минимумына сәйкес келетін полипептидтік тізбектің ең тұрақты конформациясы болып табылады;
5) L-амин қышқылдарының полипептидтік тізбегінде әдетте белоктарда кездесетін оң жақ спираль сол жаққа қарағанда әлдеқайда тұрақты.
α-спиралдың пайда болу мүмкіндігібелоктың бастапқы құрылымына байланысты. Кейбір амин қышқылдары пептидтік арқаның бұралуын болдырмайды. Мысалы, глутамат пен аспартаттың көршілес карбоксил топтары бірін-бірі кері тебеді, бұл α-спиральда сутектік байланыстың түзілуіне жол бермейді. Дәл сол себепті оң зарядталған лизин мен аргинин қалдықтары бір-біріне жақын орналасқан жерлерде тізбекті орамдау қиын. Дегенмен, пролин α-спиральді бұзуда ең үлкен рөл атқарады. Біріншіден, пролинде азот атомы қатты сақинаның бөлігі болып табылады, ол N-C байланысының айналасында айналуды болдырмайды, екіншіден, пролин азот атомында сутегінің болмауына байланысты сутектік байланыс түзбейді.
β-бүктеу – қабатты құрылымсызықты орналасқан пептидтік фрагменттердің арасындағы сутектік байланыстар арқылы түзіледі. Екі тізбек те тәуелсіз болуы мүмкін немесе бір полипептидтік молекулаға жатады. Егер тізбектер бір бағытта бағытталған болса, онда мұндай β-құрылым параллель деп аталады. Тізбектердің бағыты қарама-қарсы болған жағдайда, яғни бір тізбектің N-соңы екінші тізбектің С ұшымен сәйкес келген кезде β-құрылым антипараллель деп аталады. Энергетикалық тұрғыдан, дерлік сызықты сутегі көпірлері бар антипараллельді β-бүктелу жақсырақ.


параллельді β-бүктелу антипараллельді β-бүктелу
α-спиралдан айырмашылығысутектік байланыстармен қаныққан, β-бүктеу тізбегінің әрбір бөлімі қосымша сутектік байланыстардың түзілуі үшін ашық. Амин қышқылдарының бүйірлік радикалдары кезекпен жоғары және төмен жапырақ жазықтығына дерлік перпендикуляр бағытталған.
Пептидтік тізбек қайдаайтарлықтай тік иіледі, жиі β-ілмек кездеседі. Бұл 4 амин қышқылы қалдықтары 180 o бүгілген және бірінші және төртінші қалдықтар арасындағы бір сутегі көпірімен тұрақтандырылған қысқа фрагмент. Ірі аминқышқылдарының радикалдары β-циклдің түзілуіне кедергі жасайды, сондықтан оған көбінесе ең кішкентай амин қышқылы глицин кіреді.
Екіншіден жоғары белоктың құрылымы- бұл қайталама құрылымдарды ауыстырудың белгілі бір тәртібі. Домен белгілі бір дәрежеде құрылымдық және қызметтік автономияға ие белок молекуласының жеке бөлігі ретінде түсініледі. Енді домендер белок молекулаларының құрылымының іргелі элементтері болып саналады, ал α-спиральдар мен β-қабаттардың орналасуларының қатынасы мен табиғаты бастапқы құрылымдарды салыстырудан гөрі белок молекулаларының эволюциясын және филогенетикалық қатынастарды түсінуге көбірек мүмкіндік береді.
Эволюцияның негізгі мақсатыжаңа белоктардың құрылысы. Қаптама шарттарын қанағаттандыратын және функционалдық міндеттердің орындалуын қамтамасыз ететін осындай аминқышқылдарының реттілігін кездейсоқ синтездеудің шексіз мүмкіндігі бар. Сондықтан әртүрлі қызметтері бар, бірақ құрылымы жағынан ұқсас белоктар жиі кездеседі, сондықтан олардың бір арғы тегі болған немесе бір-бірінен дамыған сияқты көрінеді. Белгілі бір мәселені шешу қажеттілігімен бетпе-бет келген эволюция алдымен бұл үшін белоктарды жобалауды емес, бұл үшін бұрыннан қалыптасқан құрылымдарды бейімдеуді, оларды жаңа мақсаттарға бейімдеуді жөн көретін сияқты.
Жиі қайталанатын екіншіден жоғары құрылымдардың кейбір мысалдары:
1) αα' – тек α-спиральдары бар белоктар (миоглобин, гемоглобин);
2) ββ' - тек β-құрылымдары бар белоктар (иммуноглобулиндер, супероксиддисмутаза);
3) βαβ' - β-бөшкенің құрылымы, әрбір β-қабат бөшкенің ішінде орналасады және молекуланың бетінде орналасқан α-спиральмен байланысты (триозафосфоизомераза, лактатдегидрогеназа);
4) «мырыш саусақ» - 20 амин қышқылы қалдықтарынан тұратын ақуыз фрагменті, мырыш атомы екі цистеин және екі гистидин қалдықтарымен байланысады, нәтижесінде 12-ге жуық амин қышқылы қалдықтарынан тұратын «саусақ» реттегішпен байланыса алады. ДНҚ молекуласының аймақтары;
5) «лейцин сыдырма» - өзара әрекеттесетін белоктардың құрамында кемінде 4 лейцин қалдығы бар α-спиральдық аймағы бар, олар бір-бірінен 6 аминқышқылынан алшақ орналасады, яғни әрбір екінші айналымның бетінде орналасады және гидрофобты түзе алады. лейцин қалдықтарымен басқа ақуызбен байланысады. Мысалы, лейцин сыдырмаларының көмегімен күшті негізді гистонды ақуыздардың молекулалары оң зарядты жеңе отырып, кешендерге біріктірілуі мүмкін.
Ақуыздың үшінші реттік құрылымы- бұл аминқышқылдарының бүйірлік радикалдары арасындағы байланыстар арқылы тұрақтандырылған ақуыз молекуласының кеңістікте орналасуы.
Ақуыздың үшінші реттік құрылымын тұрақтандыратын байланыс түрлері:

электростатикалық сутегі гидрофобты дисульфидті әрекеттесулер байланыстар өзара әрекеттесулер байланыстар
Бүктелуге байланыстыҮшіншілік құрылымды ақуыздарды екі негізгі түрге жіктеуге болады - фибриллярлы және глобулярлы.
фибриллярлық ақуыздар- суда ерімейтін ұзын жіп тәрізді молекулалар, олардың полипептидтік тізбектері бір ось бойымен созылған. Бұл негізінен құрылымдық және жиырылғыш белоктар. Ең көп таралған фибриллярлық ақуыздардың бірнеше мысалдары:
1. α-Кератиндер. Эпидермис жасушаларымен синтезделеді. Олар шаштың, жүннің, қауырсынның, мүйіздің, тырнақтың, тырнақтың, иненің, таразының, тұяқтың және тасбақа қабығының құрғақ салмағының барлығына дерлік, сонымен қатар терінің сыртқы қабатының салмағының едәуір бөлігін құрайды. Бұл ақуыздардың тұтас отбасы, олар аминқышқылдарының құрамы бойынша ұқсас, құрамында көптеген цистеин қалдықтары бар және полипептидтік тізбектердің кеңістіктік орналасуы бірдей.
Шаш жасушаларында кератиннің полипептидтік тізбектеріалдымен талшықтарға бөлінеді, содан кейін құрылымдар арқан немесе бұралған кабель тәрізді қалыптасады, олар ақырында жасушаның бүкіл кеңістігін толтырады. Бұл кезде шаш жасушалары тегістеліп, ақырында өледі, ал жасуша қабырғалары әр шаштың айналасында кутикула деп аталатын түтік тәрізді қабық түзеді. α-кератинде полипептидтік тізбектер α-спираль түрінде болады, бірінен соң бірін айналдыра үш өзекті кабельге айналдырып, дисульфидті айқас байланыстар түзеді.
N-терминал қалдықтары орналасқанбір жағында (параллель). Кератиндер суда ерімейді, олардың құрамында полярлы емес жанама радикалдары бар аминқышқылдары басым болғандықтан, олар сулы фазаға қарай бұрылады. Пермь кезінде келесі процестер жүреді: біріншіден, дисульфидті көпірлер тиолдармен тотықсыздану арқылы жойылады, содан кейін шашқа қажетті пішін берілгенде, оны қыздыру арқылы кептіреді, ал ауа оттегімен тотығу нәтижесінде жаңа дисульфидті көпірлер пайда болады. шаш үлгісінің пішінін сақтайды.
2. β-кератиндер. Оларға жібек және өрмекші фиброин жатады. Олар құрамында глицин, аланин және серин басым болатын антипараллельді β қатпарлы қабаттар.
3. Коллаген. Жоғары сатыдағы жануарларда ең көп таралған ақуыз және дәнекер тіндердің негізгі фибриллярлық ақуызы. Коллаген фибробласттарда және хондроциттерде – арнайы дәнекер тінінің жасушаларында синтезделеді, содан кейін ол сыртқа шығарылады. Коллаген талшықтары теріде, сіңірлерде, шеміршектерде және сүйектерде кездеседі. Олар созылмайды, беріктігі бойынша болат сымнан асып түседі, коллаген фибрилдері көлденең жолақпен сипатталады.
Суда қайнатқанда талшықты, ерімейтін және сіңірілмейтін коллаген кейбір коваленттік байланыстардың гидролизі нәтижесінде желатинге айналады. Коллаген құрамында 35% глицин, 11% аланин, 21% пролин және 4-гидроксипролин (тек коллаген мен эластинде болатын амин қышқылы) бар. Бұл композиция желатиннің тағамдық ақуыз ретінде салыстырмалы түрде төмен тағамдық құндылығын анықтайды. Коллаген фибрилдері тропоколлаген деп аталатын қайталанатын полипептидтік суббірліктерден тұрады. Бұл суббірліктер фибрилдің бойымен параллель шоғырлар түрінде басынан құйрығына дейін орналасқан. Бастардың жылжуы тән көлденең жолақты береді. Бұл құрылымдағы бос орындар, қажет болған жағдайда, сүйек минерализациясында маңызды рөл атқаратын гидроксиапатит Са 5 (OH) (PO 4) 3 кристалдарының тұндыру орны бола алады.


Тропоколлаген суббірліктер болып табыладыα- және β-кератиндерден өзгеше, үш ядролы арқан түрінде тығыз бұралған үш полипептидтік тізбектен тұрады. Кейбір коллагендерде барлық үш тізбектің аминқышқылдарының реттілігі бірдей, ал басқаларында тек екі тізбек бірдей, ал үшіншісі олардан ерекшеленеді. Тропоколлаген полипептидтік тізбегі пролин мен гидроксипролиннің әсерінен тізбектің иілуіне байланысты бір айналымда тек үш аминқышқылының қалдығы болатын сол жақ спиралды құрайды. Үш тізбек сутектік байланыстардан басқа, көрші тізбектерде орналасқан екі лизин қалдығының арасында түзілетін ковалентті типті байланыс арқылы өзара байланысады:

Біз қартайған сайын, тропоколлаген бөлімшелерінің ішінде және олардың арасында кросс-байланыстардың көбеюі түзіледі, бұл коллаген фибрилдерін қаттырақ және сынғыш етеді, бұл шеміршек пен сіңірлердің механикалық қасиеттерін өзгертеді, сүйектерді сынғыш етеді және көздің мүйізді қабығының мөлдірлігін төмендетеді. көз.
4. Эластин. Байламдардың сары серпімді тінінде және ірі артериялардың қабырғаларында дәнекер тінінің серпімді қабатында болады. Эластиндік фибрилдердің негізгі суббірлігі тропоэластин болып табылады. Эластин глицин мен аланинге бай, құрамында лизин көп, пролин аз. Эластиннің бұрандалы бөліктері созылған кезде созылады, бірақ жүкті алып тастаған кезде бастапқы ұзындығына оралады. Төрт түрлі тізбектің лизин қалдықтары бір-бірімен ковалентті байланыстар құрайды және эластиннің барлық бағытта қайтымды созылуына мүмкіндік береді.
Глобулярлы белоктар- полипептидтік тізбегі ықшам глобулға бүктелген белоктар алуан түрлі функцияларды орындауға қабілетті.
Глобулярлы белоктардың үшінші реттік құрылымыонда миоглобиннің мысалын қарастыру өте ыңғайлы. Миоглобин - бұлшықет жасушаларында кездесетін салыстырмалы түрде аз мөлшерде оттегімен байланыстыратын ақуыз. Ол байланысқан оттегін сақтайды және оның митохондрияға өтуіне ықпал етеді. Миоглобин молекуласында бір полипептидтік тізбек және бір гемотоп (гем) – протопорфириннің темірмен кешені болады.
Негізгі қасиеттер миоглобин:
а) миоглобин молекуласы ықшам болғаны сонша, оның ішіне тек 4 су молекуласы сыйды;
б) барлық полярлы аминқышқылдарының қалдықтары, екеуінен басқасы, молекуланың сыртқы бетінде орналасқан және олардың барлығы гидратталған күйде болады;
в) гидрофобты амин қышқылы қалдықтарының көпшілігі миоглобин молекуласының ішінде орналасады және осылайша сумен жанасудан қорғалған;
г) миоглобин молекуласындағы төрт пролин қалдықтарының әрқайсысы полипептидтік тізбектің иілу жерінде, серин, треонин және аспарагин қалдықтары иілудің басқа жерлерінде орналасады, өйткені мұндай аминқышқылдары α-спираль түзілуіне кедергі келтіреді, егер олар бір-бірімен;
д) тегіс гемотоп молекула бетіне жақын қуыста (қалтада) жатады, темір атомының гем жазықтығына перпендикуляр бағытталған екі координациялық байланысы бар, олардың біреуі гистидин қалдығы 93пен байланысады, ал екіншісі байланыстыру қызметін атқарады. оттегі молекуласы.
Ақуыздың үшінші реттік құрылымынан басталадыөзінің биологиялық қызметін атқаруға қабілетті болады. Белоктардың жұмыс істеуі белоктың бетіне үшінші реттік құрылым төселген кезде лигандтар деп аталатын басқа молекулаларды өзіне қоса алатын орындардың пайда болуына негізделген. Ақуыздың лигандпен әрекеттесуінің жоғары ерекшелігі активті орталықтың құрылымының лиганд құрылымымен комплементарлылығымен қамтамасыз етіледі. Толықтырғыштық - өзара әрекеттесетін беттердің кеңістіктік және химиялық сәйкестігі. Көптеген белоктар үшін үшінші реттік құрылым қатпарланудың максималды деңгейі болып табылады.
Төрттік белок құрылымы- тек қана ковалентті емес байланыстармен, негізінен электростатикалық және сутегімен байланысқан екі немесе одан да көп полипептидтік тізбектерден тұратын белоктарға тән. Көбінесе белоктар екі немесе төрт суббірліктен тұрады, төрттен көп суббірліктерде әдетте реттеуші ақуыздар болады.
Төрттік құрылымы бар белоктаржиі олигомерлі деп аталады. Гомомерлі және гетеромерлі белоктарды ажыратыңыз. Гомер белоктары - барлық суббірліктер құрылымы бірдей белоктар, мысалы, каталаза ферменті төрт абсолютті бірдей суббірліктерден тұрады. Гетеромерлі белоктардың әртүрлі суббірліктері бар, мысалы, РНҚ-полимераза ферменті әртүрлі қызмет атқаратын әр түрлі құрылымдағы бес бөлімшеден тұрады.
Бір суббірліктің өзара әрекеттесуіспецификалық лигандпен бүкіл олигомерлі белоктың конформациялық өзгерістерін тудырады және лигандтарға басқа суббірліктердің жақындығын өзгертеді, бұл қасиет олигомерлі белоктардың аллостериялық реттелу қабілетінің негізінде жатыр.
Ақуыздың төрттік құрылымын қарастыруға болады b гемоглобин мысалында. Оның құрамында темір атомдары Fe 2+ темір түрінде болатын төрт полипептидтік тізбек және төрт гемдік протездік топ бар. Молекуланың белоктық бөлігі – глобин екі α-тізбектен және екі β-тізбектен тұрады, құрамында 70%-ға дейін α-спиральдары бар. Төрт тізбектің әрқайсысының өзіне тән үшінші құрылымы бар және әрбір тізбекпен бір гемотоп байланысады. Әртүрлі тізбектердің гемдері салыстырмалы түрде алыс және әртүрлі көлбеу бұрыштары бар. Екі α-тізбек пен екі β-тізбегі арасында аз тікелей байланыстар түзіледі, ал α- және β-тізбектер арасында гидрофобты радикалдармен түзілген α 1 β 1 және α 2 β 2 типті көптеген контактілер пайда болады. Арна α 1 β 1 және α 2 β 2 арасында қалады.
Миоглобиннен айырмашылығыгемоглобин сипатталадыоттегіге айтарлықтай төмен жақындық, бұл тіндерде бар оттегінің төмен парциалды қысымында оларға байланыстырылған оттегінің маңызды бөлігін беруге мүмкіндік береді. Өкпе альвеолаларына тән рН жоғары және төмен CO 2 концентрацияларында оттегі гемоглобин темірімен оңай байланысады; гемоглобиннен оттегінің бөлінуіне тіндерге тән рН төмен мәндері және CO 2 жоғары концентрациясы қолайлы.
Гемоглобин оттегімен қатар сутегі иондарын да тасымалдайды., олар тізбектердегі гистидин қалдықтарымен байланысады. Гемоглобин сонымен қатар төрт полипептидтік тізбектің әрқайсысының терминалдық амин тобына қосылатын көмірқышқыл газын тасымалдайды, нәтижесінде карбаминогемоглобин түзіледі:

INэритроциттер жеткілікті жоғары концентрацияларда 2,3-дифосфоглицерат (DFG) заты бар, оның мөлшері биіктікке көтерілген сайын және гипоксия кезінде ұлғайып, тіндердегі гемоглобиннен оттегінің босатылуын жеңілдетеді. DFG α 1 β 1 және α 2 β 2 аралығындағы арнада β-тізбектерінің оң жұқтырылған топтарымен әрекеттеседі. Оттегі гемоглобинмен байланысқанда, DPG қуыстан ығысады. Кейбір құстардың эритроциттерінде ДПГ емес, инозитол гексафосфат болады, ол гемоглобиннің оттегіге жақындығын одан әрі төмендетеді.
 2,3-дифосфоглицерат (DPG)
2,3-дифосфоглицерат (DPG)
HbA - қалыпты ересек гемоглобин, HbF – ұрықтың гемоглобині, O 2 , HbS – гемоглобинге орақ тәрізді жасушалық анемияда үлкен жақындығы бар. Орақ жасушалы анемия - гемоглобиннің генетикалық ауытқуымен байланысты ауыр тұқым қуалайтын ауру. Ауру адамдардың қанында, біріншіден, оңай жыртылатын, екіншіден, қан капиллярларын бітеп тастайтын жұқа орақ тәрізді қызыл қан жасушаларының әдеттен тыс көп саны бар.
Молекулалық деңгейде гемоглобин S ерекшеленедігемоглобин А-дан, β-тізбектерінің 6-позициясындағы бір амин қышқылы қалдығы, мұнда глутамин қышқылының қалдығының орнына валин орналасқан. Осылайша, гемоглобин S құрамында екі теріс заряд аз болады, валиннің пайда болуы молекула бетінде «жабысқақ» гидрофобты контактінің пайда болуына әкеледі, нәтижесінде деоксигенация кезінде дезоксигемоглобин S молекулалары бір-біріне жабысып, ерімейтін әдеттен тыс ұзын жіпшелерді түзеді. эритроциттердің деформациясына әкелетін агрегаттар.
Ақуыздың құрылымдық ұйымдасу деңгейінің бастапқыдан жоғары қалыптасуына тәуелсіз генетикалық бақылау бар деп ойлауға негіз жоқ, өйткені біріншілік құрылым екіншілік, үшіншілік және төрттік (бар болса) анықтайды. Белоктың нативті конформациясы берілген шарттарда ең термодинамикалық тұрақты құрылым болып табылады.
ДӘРІС 6
Белоктардың физикалық, химиялық және биологиялық қасиеттері бар.
Белоктардың физикалық қасиеттерібелоктардың сфералық емес пішініне байланысты молекулалық салмақтың болуы, қос сыну (қозғалыстағы ақуыз ерітіндісінің оптикалық сипаттамаларының тыныштықтағы ерітіндімен салыстырғанда өзгеруі), ақуыз молекулаларының зарядына байланысты электр өрісіндегі қозғалғыштығы. Сонымен қатар, белоктар оптикалық қасиеттермен сипатталады, олар жарықтың поляризация жазықтығын айналдыру, ақуыз бөлшектерінің үлкен мөлшеріне байланысты жарық сәулелерін шашырату және ультракүлгін сәулелерді сіңіру қабілетінен тұрады.
тән физикалық қасиеттердің бірібелоктар – бетінде адсорбциялану, кейде молекулалардың ішіне, төмен молекулалық органикалық қосылыстар мен иондарды ұстау қабілеті.
Белоктардың химиялық қасиеттері әртүрліерекше әртүрлілік, өйткені белоктар амин қышқылы радикалдарының барлық реакцияларымен сипатталады және пептидтік байланыстардың гидролиз реакциясы тән.
Қышқылдық және негіздік топтардың айтарлықтай санының болуыбелоктар амфотерлік қасиет көрсетеді. Бос аминқышқылдарынан айырмашылығы белоктардың қышқылдық-негіздік қасиеттері пептидтік байланысты түзуге қатысатын α-амин және α-карбоксиді топтармен емес, аминқышқылдары қалдықтарының зарядталған радикалдарымен анықталады. Белоктардың негізгі қасиеттері аргинин, лизин және гистидин қалдықтарына байланысты. Қышқылдық қасиеттері аспарагин және глутамин қышқылдарының қалдықтарына байланысты.
Ақуызды титрлеу қисықтары жеткіліктіТүсіндіру қиын, өйткені кез келген ақуыздың титрленетін топтары тым көп, белоктың иондалған топтары арасында электростатикалық әрекеттесулер болады және әрбір титрленетін топтың pK-сына көршілес гидрофобты қалдықтар мен сутектік байланыстар әсер етеді. Ең үлкен практикалық қолдану ақуыздың изоэлектрлік нүктесі болып табылады - ақуыздың жалпы заряды нөлге тең рН мәні. Изоэлектрлік нүктеде ақуыз максималды инертті, электр өрісінде қозғалмайды және ең жұқа гидратталған қабықшаға ие.
Белоктар буферлік қасиет көрсетеді, бірақ олардың буферлік сыйымдылығы шамалы. Ерекшелік - құрамында гистидин қалдықтарының көп мөлшері бар ақуыздар. Мысалы, эритроциттердің құрамындағы гемоглобин гистидин қалдықтарының өте жоғары болуына байланысты, шамамен 7 рН кезінде маңызды буферлік сыйымдылыққа ие, бұл эритроциттердің оттегі мен көмірқышқыл газын тасымалдаудағы рөлі үшін өте маңызды. қан.
Белоктар суда ериді, және физикалық тұрғыдан олар шынайы молекулалық ерітінділерді құрайды. Бірақ белок ерітінділері кейбір коллоидтық қасиеттермен сипатталады: Тендаль эффектісі (жарық шашырау құбылысы), жартылай өткізгіш мембраналардан өте алмау, жоғары тұтқырлық, гель түзілуі.
Ақуыздың ерігіштігі өте тәуелдітұздардың концентрациясы бойынша, яғни ерітіндінің иондық күші бойынша. Дистилденген суда белоктар көбінесе нашар ериді, бірақ иондық күші артқан сайын олардың ерігіштігі артады. Бұл жағдайда гидратталған бейорганикалық иондардың көбеюі белоктың бетімен байланысады және сол арқылы оның агрегация дәрежесі төмендейді. Жоғары иондық күште тұз иондары белок молекулаларынан гидратация қабығын алады, бұл ақуыздардың агрегациясына және тұнбаға түсуіне әкеледі (тұздану құбылысы). Ерігіштік айырмашылығын пайдаланып, ас тұздарының көмегімен белоктар қоспасын бөлуге болады.
Белоктардың биологиялық қасиеттерінің қатарындаең алдымен олардың каталитикалық белсенділігіне байланысты. Белоктардың тағы бір маңызды биологиялық қасиеті – олардың гормондық белсенділігі, яғни организмдегі реакциялардың барлық топтарына әсер ету қабілеті. Кейбір белоктар улы қасиеттерге, патогендік белсенділікке, қорғаныс және рецепторлық қызметтерге ие және жасушалардың адгезия құбылыстарына жауап береді.
Белоктардың тағы бір ерекше биологиялық қасиеті- денатурация. Табиғи күйіндегі белоктарды нативті белоктар деп атайды. Денатурация – денатурациялаушы заттардың әсерінен белоктардың кеңістіктік құрылымының бұзылуы. Денатурация кезінде белоктардың біріншілік құрылымы бұзылмайды, бірақ олардың биологиялық белсенділігі жойылады, сонымен қатар ерігіштік, электрофоретикалық қозғалғыштық және кейбір басқа реакциялар. Ақуыздың белсенді орталығын құрайтын аминқышқылдарының радикалдары денатурация кезінде бір-бірінен кеңістікте алшақ орналасады, яғни белоктың лигандпен байланысуының арнайы орталығы бұзылады. Әдетте глобулярлы белоктардың гидрофобты өзегінде орналасатын гидрофобты радикалдар денатурация кезінде молекуланың бетінде пайда болып, сол арқылы тұнбаға түсетін белоктардың агрегациясына жағдай жасайды.
Белоктың денатурациясын тудыратын реагенттер мен жағдайлар:
60 ° C жоғары температура - ақуыздағы әлсіз байланыстардың бұзылуы,
Қышқылдар мен сілтілер – ионогендік топтардың иондануының өзгеруі, иондық және сутектік байланыстардың үзілуі,
Мочевина - мочевинамен сутектік байланыстардың түзілуі нәтижесінде молекулаішілік сутектік байланыстардың бұзылуы,
Алкоголь, фенол, хлорамин - гидрофобты және сутегі байланыстарын бұзу,
Ауыр металл тұздары – ауыр металл иондарымен ерімейтін ақуыз тұздарының түзілуі.
Денатурациялаушы агенттерді жою арқылы ренатурация мүмкін болады, өйткені пептидтік тізбек ерітіндідегі ең аз бос энергиямен конформацияны қабылдауға бейім.
Жасуша жағдайында белоктар мүмкінжоғары температураға қарағанда баяуырақ болса да, өздігінен денатурацияланады. Жасушадағы белоктардың өздігінен регенерациялануы қиын, өйткені жоғары концентрацияға байланысты жартылай денатуратталған молекулалардың агрегациялану ықтималдығы жоғары.
Жасушаларда белоктар болады- тұрақсыз, агрегацияға бейім күйдегі ішінара денатуратталған ақуыздармен байланысу және өзінің нативті конформациясын қалпына келтіру қабілеті бар молекулалық шаперондар. Бастапқыда бұл белоктар жылу соққысының белоктары ретінде ашылды, өйткені олардың синтезі жасушаға стресстік әсер ету кезінде, мысалы, температураның жоғарылауымен күшейтілді. Шаперондар суббірліктердің массасына қарай жіктеледі: hsp-60, hsp-70 және hsp-90. Әрбір класқа бір-бірімен байланысты ақуыздар тобы кіреді.
Молекулярлық шаперондар ( hsp-70)жасушаның барлық бөліктерінде: цитоплазмада, ядрода, эндоплазмалық ретикулумда, митохондрияда кездесетін жоғары дәрежеде сақталған белоктар класы. Бір полипептидтік тізбектің С-соңында hsp-70 гидрофобты радикалдармен байытылған ұзындығы 7-9 аминқышқылының қалдығы бар пептидтермен әрекеттесе алатын ойық болып табылатын аймаққа ие. Глобулярлы ақуыздардағы мұндай орындар шамамен әрбір 16 аминқышқылында кездеседі. Hsp-70 ақуыздарды термиялық инактивациядан қорғауға және ішінара денатуратталған ақуыздардың конформациясы мен белсенділігін қалпына келтіруге қабілетті.
Шаперондар-60 (hsp-60)белоктардың үшінші реттік құрылымын құруға қатысады. Hsp-60 14 суббірліктен тұратын олигомерлік ақуыздар ретінде қызмет етеді. Hsp-60 екі сақина құрайды, әрбір сақина бір-бірімен байланысқан 7 суббірліктен тұрады.
Әрбір ішкі бірлік үш доменнен тұрады:
Апикальды доменде суббірліктерден құралған қуыстың ішіне қарайтын бірқатар гидрофобты аминқышқылдарының қалдықтары бар;
Экваторлық домен АТФаза белсенділігіне ие және шаперонин кешенінен ақуызды босату үшін қажет;
Аралық домен апикальды және экваторлық домендерді байланыстырады.
Бетінде фрагменттері бар белокгидрофобты аминқышқылдарымен байытылған шаперонин кешенінің қуысына енеді. Бұл қуыстың спецификалық ортасында, жасуша цитозолының басқа молекулаларынан оқшауланған жағдайда, мүмкін белоктық конформацияларды таңдау энергетикалық қолайлы конформация табылмайынша жүреді. Нәтижелі конформацияның шаперонға тәуелді қалыптасуы энергияның айтарлықтай көлемін тұтынумен байланысты, оның көзі АТФ болып табылады.
Белоктар (белоктар) – пептидтік табиғаттың жоғары молекулалы полимерлі қосылыстары (полигетерамин қышқылдары).
Белоктардың біріншілік құрылымы – полипептидтік тізбектегі (ППК) аминқышқылдары қалдықтарының кезектесуі.
Белоктардың негізгі құрылымы коваленттіқұрылымы, өйткені ол негізделген пептидаминқышқылдарының а-амин және а-карбоксил топтары арасындағы байланыс. Нәтижесінде полипептидтік тізбектер тармақталмаған.
Полипептидтік тізбектің қаңқасы (омыртқасы, қаңқасы) үнемі қайталанатын құрылымдық элементтерден тұрады.
Полипептидтік тізбектің векторы бар, тізбектің бағыты N-соңғысынан (тізбектің басы) С-соңына (тізбектің соңы), N-соңы - бос а болатын ұшы. -амин тобы орналасқан. С-соңы - бос а-карбоксил тобы орналасқан соң. Белоктардың аминқышқылдарының тізбегі N-соңынан бастап үш әріпті аминқышқылдарының аббревиатуралары арқылы белгіленеді, мысалы: gli-ala-cis-pro. Ақуыздағы амин қышқылы қалдықтарының бір әріптік белгісін де қолдануға болады.
Белоктардың N- және С-терминалдарын өзгертуге болады. N-соңғы жағындағы амин тобы ацетилденген, формилденген немесе метилденген болуы мүмкін. Бірқатар ақуыздарда N-терминал бос амин тобын қамтымайтын пирролидон карбонатының (пироглутамат) қалдығы болып табылады. С-терминусы амидациялануы мүмкін. C-терминал модификациялары N-терминал модификацияларына қарағанда сирек кездеседі.
Ақуыздың поликонденсация коэффициенті 50-ден 2500-ге дейін ауытқиды. Әдетте белоктың құрамында 100-300 аминқышқылының қалдықтары болады. Бір амин қышқылы қалдығының орташа молекулалық салмағы шамамен 110 Да болғандықтан, белоктардың молекулалық салмағы 6000-нан миллиондаған Да-ға дейін өзгереді.
Әрбір жеке ақуыздың бірегей бастапқы құрылымы бар. Алғашқы құрылымы анықталған белок инсулин болды. Сангер жасады. Оның стратегиясы келесідей болды. Ол алдымен екі полипептидтік тізбекті бөліп алды, содан кейін олардың арнайы ферментативті бөлінуін бір-бірінен тұратын тізбектері бар шағын пептидтерге жүргізді. Содан кейін 1-фтор-2,4-динитробензолдың көмегімен N-терминал қалдықтары анықталды. Сонымен қатар, ол пептидтердің аминқышқылдық құрамын анықтады және соңында пептидтердің қабаттасу ретін салыстыру арқылы олардың құрылымын анықтай алды. Жалпы алғанда, Сэнгердің стратегиясы бүгінгі күнге дейін өзінің маңыздылығын сақтап қалды. Дегенмен, басқа тәсілдер де ұсынылды. Эдман N-терминал аминқышқылдарының қалдықтарын дәйекті бөлу және идентификациялаудың автоматты процедурасының әдісін жасады. Бастапқы құрылымды ашу үшін рентгендік дифракциялық талдауды қолдануға болады. Амин қышқылы қалдықтарының ретін хабаршы РНҚ-ның нуклеотидтер тізбегінен анықтауға болады.
Қазіргі уақытта 2000-нан астам белоктардың бастапқы құрылымы анықталды. Теориялық тұрғыдан белоктардың біріншілік құрылымының әртүрлі нұсқаларының саны шексіз. Тіпті 20 түрлі аминқышқылдарынан тұратын полипептид үшін ықтимал тізбектердің саны 20×10 18 құрайды. Тірі табиғатта мүмкін болатын тізбектердің аз ғана бөлігі ғана жүзеге асады, олардың жалпы саны тірі ағзалардың барлық түрлерінде 10 10 -10 12. шамасында бағаланады.
Белоктардың бастапқы құрылымы генетикалық түрде анықталады, яғни. ақуыздағы аминқышқылдарының реттілігі ондағы нуклеотидтердің реттілігімен анықталады ДНҚ. ДНҚ нуклеотидтер тізбегінің бұрмалануы молекулалық патологияның себебі болып табылатын биологиялық қасиеттері өзгерген аномальды белоктардың пайда болуына әкеледі. Атап айтқанда, орақ жасушалы анемияның себебі гемоглобиннің b-тізбегін бақылайтын геннің нүктелік мутациясы болып табылады. Мұның салдары b-тізбегінің 6-шы позициясындағы глутамат қалдығының валинмен алмастырылуы болып табылады. Мұндай алмастыру екі b-жіптің әрқайсысында бір теріс зарядтың жоғалуына әкеледі, бұл гемоглобиннің конформациясының өзгеруіне және оның биологиялық функциясының жоғалуына әкеледі.
Гомологиялық белоктар – әртүрлі түрлерде бірдей қызмет атқаратын белоктар. Мысал ретінде гемоглобинді келтіруге болады: барлық омыртқалы жануарларда ол оттегін тасымалдаумен байланысты бірдей функцияны орындайды. Гомологиялық белоктар бірдей аминқышқылдарының көптеген позицияларда болуымен сипатталады. Белгілі болғандай, гомологтық белоктар ерекшеленетін аминқышқылдарының қалдықтарының саны осы түрлер арасындағы филогенетикалық айырмашылыққа пропорционалды. Мысалы, жылқы мен ашытқы С цитохромының молекулалары 48 аминқышқылының қалдықтарымен ерекшеленеді, ал бірдей тауық пен үйрек молекулалары тек 2 қалдықпен ерекшеленеді. Тауық пен күркетауықтың цитохром Cs-іне келетін болсақ, олардың аминқышқылдарының тізбегі бірдей. Әртүрлі түрлердің гомологтық белоктарының аминқышқылдарының тізбектеріндегі айырмашылықтар саны туралы мәліметтер эволюциялық процесте әртүрлі жануарлар мен өсімдіктер түрлерінің пайда болуы мен дамуының кезекті кезеңдерін көрсететін эволюциялық карталарды құру үшін пайдаланылады.




