Millä levillä ei ole siima-asteita. Alemmat kasvit tai levät. Yksinäiset vapaasti elävät muodot
Monadiset vegetatiiviset solut ja levien elinkaaren monadiset vaiheet (eläinporit ja sukusolut) on varustettu flagellalla - pitkillä ja melko paksuilla solujen kasvaimilla, jotka on ulkopuolelta peitetty plasmalemmalla. Niiden lukumäärä, pituus, morfologia, kiinnittymispaikka ja liikelaji ovat levissä varsin erilaisia, mutta sukulaisryhmissä vakioita.
Flagella voi olla kiinnittynyt solun etupäähän (apikaalisesti) tai se voi olla hieman siirtynyt sivulle (subapaisesti); niiden kiinnittyminen on mahdollista solun sivulla (lateraalisesti) ja solun ventraalisella puolella (ventraalisesti). Morfologialtaan identtisiä lippuja kutsutaan isomorfinen jos ne eroavat - heteromorfinen. Isocont- nämä ovat samanpituisia lippuja, heterokontakti- eri pituuksia.
Flagellalla on yksi rakennesuunnitelma. On mahdollista erottaa vapaa osa (undulipodia), siirtymäalue, perusrunko (kinetosomi). Siiman eri osat eroavat toisistaan luurangon muodostavien mikrotubulusten lukumäärän ja järjestelyn suhteen (kuva 2).
Riisi. Kuva 2. Kaavio leväsiimojen rakenteesta (luokituksen: L.L. Velikanov et ai., 1981): 1 - siimojen pituusleikkaus; 2, 3 - poikittaisleikkaus flagellumin kärjen läpi; 4 - poikkileikkaus undulipodian läpi; 5 - siirtymävyöhyke; 6 - poikkileikkaus flagellumin pohjan läpi - kinetosomi
Undulipodium(käännetty latinan sanasta "waveleg") pystyy suorittamaan rytmisiä aaltomaisia liikkeitä. Undulipodium on kalvolla päällystetty aksoneemi. aksoneemi koostuu yhdeksästä ympyrän muotoon järjestetyn mikrotubulusparista ja keskellä olevasta mikrotubulusparista (kuva 2). Flagella voi olla sileä tai peitetty suomuilla tai mastigonemeilla (karvat), kun taas dinofyyteissä ja kryptofyyteissä ne ovat sekä suomujen että karvojen peitossa. Primesiofyyttien, kryptofyyttien ja viherlevien liput voidaan peittää erimuotoisilla ja -kokoisilla suomuilla.
siirtymäalue. Sillä on toiminnallisesti rooli siiman vahvistamisessa sen poistumiskohdassa solusta. Levissä erotetaan useita tyyppejä siirtymävyöhykkeen rakenteita: poikittaislevy (dinofyytit), tähtirakenne (vihreä), siirtymäspiraali (heterokontti), siirtymäsylinteri (primnesiofyytit ja dinofyytit).
Perusrunko tai kinetosomi. Tällä flagellumin osalla on rakenne onton sylinterin muodossa, jonka seinämän muodostaa yhdeksän mikrotubulusten kolmikkoa. Kinetosomin tehtävänä on yhdistää siima solun plasmalemmaan. Useiden levien tyvikappaleet voivat osallistua ydinfissioon ja niistä voi tulla mikrotubulusten organisoinnin keskuksia.
Työ loppu -
Tämä aihe kuuluu:
Merilevä
Kalastusyliopisto. Meribiologian instituutti nimetty v Zhirmunsky Dvorin mukaan.
Jos tarvitset lisämateriaalia tästä aiheesta tai et löytänyt etsimääsi, suosittelemme käyttämään hakua teostietokannassamme:
Mitä teemme saadulla materiaalilla:
Jos tämä materiaali osoittautui hyödylliseksi sinulle, voit tallentaa sen sivullesi sosiaalisissa verkostoissa:
| twiittaa |
Kaikki tämän osion aiheet:
Solujen kannet
Solupäällysteet antavat solujen sisäisen sisällön vastustuskyvyn ulkoisille vaikutuksille ja antavat soluille tietyn muodon. Kannet läpäisevät vettä ja siihen liuenneita pieniä molekyylejä
Mitokondriot
Mitokondrioita löytyy eukaryoottisten levien soluista. Leväsolujen mitokondrioiden muoto ja rakenne ovat monimuotoisempia kuin korkeampien kasvien mitokondrioissa. Ne voivat olla pyöreitä.
Pigmentit
Kaikki levät erottuvat hyvin fotosynteettisten pigmenttien joukosta. Tällaisilla ryhmillä kasvien taksonomiassa on osastojen asema. Kaikkien levien pääpigmentti on vihreä pigmentti.
plastidit
Eukaryoottisten levien solujen pigmentit sijaitsevat plastideissa, kuten kaikissa kasveissa. Levissä on kahta tyyppiä plastideja: värilliset kloroplastit (kromatoforit) ja värittömät leukoplastit (ami
Ydin ja mitoosilaitteisto
Levän ytimellä on eukaryooteille tyypillinen rakenne. Tumien lukumäärä solussa voi vaihdella yhdestä useaan. Ulkopuolella ydin on peitetty kalvolla, joka koostuu kahdesta kalvosta, ulkokalvosta
Monadinen (liputettu) tallusrakenne
Tyypillisin tämän tyyppisen rakenteen määräävä piirre on siipien esiintyminen, joiden avulla monadiset organismit liikkuvat aktiivisesti vesiympäristössä (kuva 9, A). Siirrettävä w
Rhizopodiaalinen (amoeboidi) rakenne
Ameboidityyppisen rakenteen merkittävimmät piirteet ovat vahvojen solupeitteiden puuttuminen ja kyky ameboidiliikenteeseen tilapäisesti solun pintaan muodostuvan qin avulla.
Palmelloid (hemimonas) -tyyppinen rakenne
Tämän tyyppiselle rakenteelle on ominaista liikkumattoman kasvin elämäntavan yhdistelmä monadisille organismeille tyypillisten soluorganellien läsnäoloon: supistuvat vakuolit, stigma, kiriste
Coccoid-rakenteen tyyppi
Tämä tyyppi yhdistää yksisoluiset ja siirtomaalevät, jotka ovat liikkumattomia kasvullisessa tilassa. Kokkoidityyppiset solut on peitetty kalvolla ja niillä on kasvityyppinen protoplasti (tonoplast ilman sokratteja).
Trichaly (filamenttimainen) tyyppinen rakenne
Rihmamaisen rakenteen tyypillinen piirre on liikkumattomien solujen rihmamainen sijoittuminen, jotka muodostuvat vegetatiivisesti solunjakautumisen seurauksena, mikä tapahtuu pääasiassa
Heterotrikaalinen (monilankainen) rakenne
Monisäikeinen tyyppi syntyi filamenttityypin perusteella. Monisäikeinen tallus koostuu enimmäkseen alustaa pitkin leviävistä vaakasuuntaisista langoista, jotka suorittavat kiinnitystehtävän, ja pystysuuntaisista, pitkin alustaa.
Parenkymaalinen (kudos) tyyppinen rakenne
Yksi multifilamenttisen talluksen evoluution suunnasta liittyi parenkymaalisen tallin syntymiseen. Solujen rajaton kasvu ja jakautuminen eri suuntiin johti muodostumiseen
Sifonityyppinen rakenne
Sifonaaliselle (ei-soluiselle) rakenteelle on ominaista talluksen puuttuminen sisällä, joka saavuttaa suhteellisen suuria, yleensä makroskooppisia kokoja ja tietyn erilaistumisasteen.
Lappopäällysteinen rakennetyyppi
Sifonipäällysteisen rakenteen pääominaisuus on kyky muodostaa monimutkaista tallia primaarisesta ei-soluisesta talluksesta, joka koostuu primäärisistä moniytimistä segmenteistä. SISÄÄN
suvuton lisääntyminen
Levien aseksuaalinen lisääntyminen suoritetaan erikoistuneiden solujen - itiöiden - avulla. Itiöitymiseen liittyy yleensä protoplastin jakautuminen osiin ja fissiotuotteiden vapautuminen niistä
yksinkertainen jako
Tämä lisääntymismenetelmä löytyy vain yksisoluisista levien muodoista. Yksinkertaisin jakautuminen tapahtuu soluissa, joilla on ameboidityyppinen kehon rakenne. Ameboidimuotojen jakautuminen
Pirstoutuminen
Fragmentoituminen on luontaista kaikille monisoluisille leväryhmille ja ilmenee erilaisia muotoja: hormogonian muodostuminen, talluksen irronneiden osien uusiutuminen, spontaani oksien putoaminen, uudelleenkasvu
Lisääntyminen versoilla, stoloneilla, hauteensilmuilla, kyhmyillä, akinetilla
Vihreiden, ruskeiden ja punaisten levien kudosmuodoissa kasvullinen lisääntyminen saa täydellisen muotonsa, joka poikkeaa vähän korkeampien kasvien vegetatiivisesta lisääntymisestä. Tiellä pitäminen
sukupuolinen lisääntyminen
sukupuolinen lisääntyminen levissä se liittyy seksuaaliseen prosessiin, joka koostuu kahden solun fuusiosta, mikä johtaa tsygootin muodostumiseen, joka kasvaa uudeksi yksilöksi tai tuottaa zoosporeja.
Ydinvaiheiden muutos
Seksuaalisen prosessin aikana sukusolujen ja niiden ytimien fuusion seurauksena ytimessä olevien kromosomien määrä kaksinkertaistuu. Tietyssä kehityssyklin vaiheessa, meioosin aikana, kromosomien lukumäärä vähenee, minkä seurauksena
Endofyytit / endotsoiitit tai endosymbiontit
Endosymbiontit tai solunsisäiset symbiontit ovat leviä, jotka elävät muiden organismien (selkärangattomien tai levien) kudoksissa tai soluissa. Ne muodostavat eräänlaisen ekologisen ryhmän.
Sinilevien (syanobakteerien) laitos - cyanophyta
Osaston nimi (kreikkalaisesta syanoksista - sininen) heijastaa näiden levien ominaista piirrettä - talluksen väriä, joka liittyy suhteellisen korkeaan sinisen pigmentin fykosyaniinipitoisuuteen. Syaani
Tilaa - krookokit
Ne esiintyvät yksisoluisina "yksinkertaisina" yksilöinä tai muodostavat useammin limakalvopesäkkeitä. Kun solut jakautuvat kahteen tasoon, muodostuu yksikerroksisia lamellipesäkkeitä. Jako kolmeen n
Punalevien laitos - rhodophyta
Osaston nimi on peräisin Kreikan sana rhodon ("radon") - vaaleanpunainen. Punalevien väri johtuu erilaisesta pigmenttien yhdistelmästä. Hän tulee harmaasta ja violetista
Tilaa Bangiaceae–Bangiales
Porphyra-suvun talli on ohuen kiiltävän levyn muodossa, jossa on sileät tai taitetut reunat ja joka koostuu yhdestä tai kahdesta kerroksesta tiukasti toisiinsa liittyviä soluja. Levyn pohja menee yleensä sisään
Tilaa Rhodimeniales - Rhodymeniales
Sparlingia-suku (Rhodimenia) - litteät levyt, joiden korkeus on enintään 45 cm, lehden ja kiilan muotoinen, laajennettu ja kämmenellä leikattu ylhäältä, vaaleanpunaisesta tai vaalean oranssista
Tilaa Coralline - Corallinales
Corallina-suku - nivelletyt, viuhkamaiset haaroittuneet pensaat, joiden korkeus on enintään 10 cm, haarautunut, kalkkipitoinen, vaaleanpunaisesta-lilasta melkein valkoiseen. Lisääntyy aseksuaalisesti ja seksuaalisesti. Spo
Gigartiinitilaus - Gigartinales
Rod Chondrus - tiheät nahkaiset rustoiset pensaat, joiden korkeus on enintään 20 cm, 3-4 kertaa haarautunut, vaaleankeltainen, vaaleanpunainen, violetti-tummanpunainen. Kasvaa rannikon alaosassa ja
Tilaa Ceramiaceae - Ceramiales
Suku Ceramium - pehmeät, pörröiset, nivelletyt pensaat, joiden korkeus on enintään 10 cm, kaksijakoisesti tai vuorotellen haarautunut, tummankeltainen, vaaleanpunainen sävy. Kahdesta neljään tilaukseen haarautuminen, rajallinen
Piilevien laitos - bacillariophyta
Osastoa kutsutaan diatomeiksi (kreikan sanasta di - kaksi, tome - leikkaus, dissekti) tai Bacillaria (bacillum - tikku). Sisältää yksisoluisen yksinäisen tai siirtomaajärjestön
Heterokont (multi-flagellate) levien laitos - heterokontophyta
Kaikilla heterokonteilla näyttää olevan flagellaarinen laite. Siimaloita on 2, ja toisessa niistä on erittäin tyypillisiä putkimaisia kolmijäsenisiä siipiä, eli karvoja - mastigonemeja. Se on käteistä
Systematiikka
Fossiiliset kokkoliitit tunnetaan mesozoisista esiintymistä, ja niitä oli runsaasti suuressa osassa juraa ja liitukaudella. Primesiofyytit saavuttivat suurimman monimuotoisuutensa myöhäisliitukaudella,
Kryptofyyttilevien (cryptomonas) laitos - cryptophyta
Laitos on nimetty tyyppisuvun Cryptomonas mukaan (kreikan sanasta kryptos - piilotettu, monas - yksilö). Sisältää yksisoluisia, liikkuvia, monadisia organismeja. Kryptofyyttisolut
A B C D E
Riisi. 53. Ulkomuoto kryptofyyttilevät (G.A. Belyakova et al., 2006 mukaan): A - Rhodomonas, B - Chroomonas, C - Cryptomonas, D - Hylomonas, E - Goniomonas voi muodostua
Viherlevien laitos - chlorophyta
Vihreälevä on laajin kaikista leväryhmistä, ja niitä on eri arvioiden mukaan 4-13-20 tuhatta lajia. Niissä kaikissa on thallin vihreä väri, mikä johtuu kloorin hallitsevuudesta
Tilaa Ulotrix - Ulotrichales
Ulotrix-suku (kuva 54). Ulotrix-lajit elävät useammin makeassa vedessä, harvemmin meri-, murtovesistöissä ja maaperässä. Ne kiinnittyvät vedenalaisiin esineisiin muodostaen kirkkaan vihreitä pensaita.
Bryopsid-järjestys – Bryopsidales
Suurin osa lajeista tavataan makeissa ja murtovesissä. Jotkut niistä kasvavat maaperällä, kivillä, hiekalla ja joskus suolamailla. Suku Briopsis - filiform pensaat jopa 6-8 s
Tilaa Volvox - Volvocales
Chlamydomonas-sukuun (kuva 57) kuuluu yli 500 yksisoluista levälajia, jotka elävät tuoreissa, matalissa, hyvin lämmitetyissä ja saastuneissa vesistöissä: lammissa, lätäköissä, ojissa jne. Jne
Osasto Charophytes (Characeae) - Charophyta
Charophytes - linja makean veden viherleviä, joka johti korkeampia kasveja. Nämä ovat muotoja, joissa on pääasiassa rihmamaista tallia. Usein tallus on pystysuora, leikattu ja kantaa ympäriinsä
Dinofyyttien (dinoflagellaattien) osasto - dinophyta
1. Osaston nimi tulee kreikan kielestä. dineo - pyöritä. Se yhdistää pääasiassa yksisoluisia monadeja, harvemmin kokkoideja, ameboideja tai palmelloideja, joskus siirtomaalaisia
Division euglenozoa - euglenoids
Nimen jako tyyppisuvun mukaan on Euglena (kreikasta eu - hyvin kehittynyt, glene - pupilli, silmä). Yhdistää yksittäisiä monadisia tai ameboideja. Harvoin löytyy
Termien sanasto
Autogamia - seksuaalinen lisääntyminen, jossa kaksi sisarhaploidista ydintä sulautuvat yhteiseen sytoplasmaan. Autospore on suvuttoman lisääntymisen rakenne, joka on
Kaikilla heterokonteilla näyttää olevan flagellaarinen laite. Siimaloita on 2, ja toisessa niistä on erittäin tyypillisiä putkimaisia kolmijäsenisiä siipiä, eli karvoja - mastigonemeja. Kahden epätasa-arvoisen siimon läsnäolo oli perustana tälle laajalle vesieliöryhmälle. monilipuinenlevät, tai heterokontami. Pituuserojen lisäksi siima eroaa myös morfologisesti: pääsiima koostuu akselista ja sen päällä olevista värekarvoista, sivusiima on sileä ja siimamainen. Pigmenttien koostumus on myös samanlainen - klorofyllit A Ja Kanssa ja lukuisia karotenoideja sekä kloroplastien rakennetta. Kloroplasteja ympäröi neljä kalvoa, joista kaksi ulkokalvoa on liitetty solun endoplasmiseen retikulumiin.
Osana Heterocont (Diversified) leväosastoa useita leväryhmiä on yhdistetty luokkiin: Brown, Golden, Yellow-Green, Rafidophytes, Sinura, Feotamnia, Eustigma, joita pidettiin aiemmin itsenäisinä osastoina.
Luokka Ruskealevä– Phaeophyceae (Fucophyceae)
luokan nimi " ruskolevät"heijastaa heidän talluksensa väriä (kreikasta. Rhaios- tumma). Väri johtuu suuresta määrästä ruskeita ja keltaisia pigmenttejä - klorofyllien lisäksi A Ja Kanssa Ja β -karoteeni niiden kloroplasteissa on ylimäärä ksantofyllejä, erityisesti fukoksantiinia.
Monisoluiset, pääasiassa suuret levät, erityisen laajalle levinneitä molempien pallonpuoliskojen kylmissä vesissä. Ruskealevistä ei tunneta yksisoluisia eikä siirtomaamuotoja. Noin 1,8 tuhatta lajia tunnetaan.
Solun rakenne. Ruskealevälajin solu on eukaryoottinen: se on muodostanut organelleja, joissa on omat kalvonsa: ydin, mitokondriot, plastidit jne. Vihreät pigmentit - klorofyllit A Ja Kanssa- peitetty lisäaineilla: oranssit karoteenit ja kellanruskeat ksantofyllit; jälkimmäisistä fukoksantiini on hallitseva. Ruskealevien soluista löytyy plastideja yhdestä useaan. Ulomman solukalvon päälle ruskealeväsolut kehittävät yleensä jäykän soluseinän.
Ydin on useimmiten yksi, mutta joskus ytimiä on useita. Joillakin edustajilla ytimien lukumäärä kasvaa solun iän kasvaessa. Ytimessä on aina yksi ydin.
Ruskealevien vegetatiivisten solujen kloroplasteissa on silmä (stigma) ja pyrenoidit, joita voi olla yksi tai useampi; joistakin lajeista puuttuu pyrenoidit. 30–40 lipidipallosta koostuva silmäsolu toimii linssinä, joka kohdistaa valon siimaturvotukseen, joka on fotoreseptori. Ruskeiden levien eläinporeilla ja sukusoluilla ei yleensä ole silmää. Vara-aineet kerrostuvat pyrenoideihin. Kloroplastit ovat useimmiten pieniä, levymäisiä, parietaalisia. Niiden muoto voi olla tähtimainen, nauhamainen tai lamellimainen. Ruskealeväsolujen kloroplastien muoto voi muuttua solun ikääntyessä. Kloroplastivaippa koostuu neljästä kalvosta, joista kaksi ulkokalvoa liittyy ER:ään. - endoplasminen verkkokalvo soluja.
Ruskealevien soluseinän muodostava selluloosa - jota joskus kutsutaan algulloosiksi - eroaa ominaisuuksiltaan korkeampien kasvien selluloosasta. Selluloosa seinämän ulkopuolella on erittäin limainen pektiinikerros, joka muodostuu pääasiassa algiinihaposta ja sen suoloista ja proteiiniyhdisteistä. Algiinihappo löytyy myös vain ruskeista levistä.
Yleiset luonteenpiirteet.
Kaikki ruskeat levät ovat fototrofit. Fotosynteesituotteet - spesifinen hiilihydraatti krysolaminariini,alkoholi mannitoli Ja lipidit. Lipidit kerääntyvät öljypisaroiden muodossa. Krysolaminariini kerrostuu kloroplastin ulkopuolelle: se sijaitsee erityisissä tyhjiöissä sytoplasmassa lähellä pyrenoidia. Mannitoli suorittaa reservin sekä osmoregulatorisen toiminnon; sen pitoisuus solun sisällä riippuu veden suolapitoisuuden muutoksista.
Ruskealevien nuoret solut sisältävät pieniä ja lukuisia tyhjiöitä, jotka kasvavat iän myötä fuusion seurauksena. Sytoplasmassa on erityisiä muodostumia - fysodeja, jotka sisältävät florotaniinit. Nämä aineet tunnetaan vain ruskealevälajeissa. Florotaniinien toiminta voi liittyä leväthallin suojaamiseen eläinten, kuten nilviäisten, syömiltä. Ruskeiden levien florotaniinit todennäköisesti estävät epifyyttisten levien ja eläinten asettumista talluksen pinnalle. Florotaniinien oletetaan osallistuvan suojautumiseen säteilyvaurioilta ja valikoivaan absorptioon vesiympäristö raskasmetalli-ioneja. Värittömät florotaniinit hapettuvat ilmassa pigmentiksi fykofeiiniksi, joka antaa kuivatuille ruskeille leville niiden tyypillisen tumman värin.
Talluksen rakenne. Ruskealevälajeissa ei ole yksisoluisia ja siirtomaamuotoja, kaikki yksilöt ovat monisoluisia. Ruskeiden levien tallin muoto on monipuolisin. Nämä ovat langat, hiipivät tai pystysuorassa seisovat, yksiriviset tai moniriviset; massiiviset kiinteät tai kaakeloidut kuoret ja kuoret; ohuet limakalvot tai tiheät nahkaiset levyt varressa, yksinkertaiset tai leikatut; kuplat ja pussit; putket ja johdot; vaihtelevasti haarautuneita ohuita pensaita tai voimakkaita pensaita. Thallin kiinnitys suoritetaan risoidien tai pohjien avulla. Pitämään ne pystyasennossa suuret pohjaeläimet muodostavat ilmakuplia, jotka ovat täynnä kaasua (kuva 29).
Riisi. 29. Ruskealevien ulkonäkö: 1 - Fucus; 2 - sakkariini; 3 - Alaria; 4 - Sargassum
Laminaria- ja Fucus-lahkojen edustajien tallit ovat ruskealevälajeista monimutkaisimmat. Niissä on merkkejä kudosten erilaistumisesta tallussolujen erikoistumisesta (kuva 30). Heidän thallissaan voidaan erottaa haukkua joka koostuu voimakkaasti värjäytyneistä soluista ja ydin, joka koostuu värittömistä soluista, jotka kerätään usein filamenteiksi. Laminariassa ytimeen muodostuu myös seulaputkia ja putkimaisia filamentteja. Ydin ei suorita vain kuljetustoimintoa, vaan myös mekaanista, koska se sisältää kierteitä, joissa on paksut pituussuuntaiset seinämät. Monien ruskealevien kuoren ja ytimen välissä voi olla suurien värittömien solujen välikerros.
Korkeus ruskealevien edustajien talli on useimmiten interkalaarinen (intercalary) ja apikaalinen (apikaalinen), harvoin basaali. Interkalaarinen kasvu voi olla diffuusia tai siinä voi olla kasvuvyöhyke. Suurissa muodoissa interkalaarinen (interkalaarinen) meristeemi tai aktiivisesti jakautuva soluryhmä sijaitsee kohdassa, jossa "lehtilehti" siirtyy "lehti" -terään. Suurilla levillä on myös meristaattinen vyöhyke talluksen pinnalla, niin kutsuttu meristodermi (eräänlainen korkeampien kasvien kambiumin analogi).
jäljentäminen ruskealevissä, kasvullisissa, aseksuaalisissa ja seksuaalisissa. Sukusolut muodostuvat monisoluisissa gametangioissa, zoosporit - sisään 
Riisi. 30. Sisäinen rakenne Laminaria(mukaan: L.L. Velikanov et al., 1991): A- varren pituusleikkaus; B- varren poikkileikkaus; SISÄÄN- pala lautasesta, jossa on zoosporangiaa
yksisilmäiset itiöt.
Vegetatiivinen lisääntyminen useissa ruskealevälajeissa voidaan suorittaa talluspaikoilla; lajeissa kiltti Sphacelaria- sikiön oksat (pesimäsilmut); suvun lajeissa Fucus- pohjassa oleva soluryhmä, joka pystyy erilaistumaan uudeksi tallukseksi.
Ruskealevien epäseksuaalinen lisääntyminen tapahtuu yleensä zoosporien avulla. Jotkut lajit lisääntyvät liikkumattomilla tetrasporeilla tai monosporeilla. Aseksuaalisen lisääntymisen itiöt muodostuvat meioosin ja sitä seuraavien mitoosien seurauksena yksisilmäiset itiöt. Seksuaalinen prosessi on isogamia, heterogamia ja oogamia. Sukusolut muodostuvat sisään monitahoinennämä gametangia- erityiset soluryhmät. Jokaiseen gametangiumin "pesään" muodostuu yksi sukusolu.
Ruskealevälle, sukupuoliferomonit- liukoiset aineet, jotka koordinoivat solujen toimintaa lisääntymisen aikana. Feromonit, joista tunnetaan vähintään kymmenen, voivat joko stimuloida anteridiajen avautumista tai houkutella urospuolisia sukusoluja naispuolisiin sukusoluihin. He ovat myös mukana lajien eristämisessä. Uskotaan, että feromoneja muodostuu useimmissa ruskealevälajeissa, joilla on seksuaalinen prosessi.
Elinkaari. Ruskeilla levillä on kaksi päätyyppiä elinkaaret. Yksi tyyppi on haplodiplobiont. Sporofyytit ovat diploideja, ts. Kaikki solut sisältävät täydellisen sarjan kromosomeja. Suvuttoman lisääntymisen itiöt muodostuvat yksisilmäisissä itiöissä. Itiöiden muodostumisen aikana tapahtuu pelkistysjako - meioosi. Tällaista pelkistystä kutsutaan itiöiseksi. Meioosin jälkeen muodostuneet solut (itiöt) ovat haploideja; sisältää puolet kromosomien määrästä. Haploidiset zoosporit ja tetrasporit itävät haploidiksi gametofyytiksi, jolle muodostuu sukusoluja monisoluisissa gametangioissa. Sukusolujen fuusion jälkeen muodostunut diploidinen tsygootti kasvaa diploidiseksi sporofyyttiksi.
Toinen tyyppi on diplobiont; sporofyytit, itiöt ja gametofyytit ovat diploideja; väheneminen jakautuu sukusolujen muodostumisen aikana. Tällaista vähennystä kutsutaan gameettiseksi. Haploidisten sukusolujen fuusion jälkeen muodostuu diploidinen tsygootti, joka itää diploidiseksi sporofyytiksi. Tämä tyyppi on yleinen rakkolevälajeissa (kuva 31).

Riisi. 31. Elinkaari Sakariinit(mukaan: L.L. Velikanov et al., 1991): 1 – sporofyytti, 2 – zoosporangian sori, 3 – eläinporofyytti , 4 - gametofyytti, 5 - anteridium, 6 - oogonium, 7 - anterotsoidi, 8 - tsygootti (hedelmöitetty muna)
Makroskooppisella sporofyytillä Sakariinit, levyn yläosaan kehittyy raidat sori(ryhmät) zoosporangia. Zoosporangioissa muodostuu pelkistysjakautumisen jälkeen zoosporeja, joissa on kaksi eriarvoista siimoa. Ne kasvavat mikroskooppisiksi gametofyyteiksi: uroksilla on anteridia ja naaraalla oogonia. Anterozoidien hedelmöittynyt munasolu itää välittömästi, ilman lepoaikaa ja muodostaa sporofyytin. Tällä elämänkaarella Sakariinit päättyy.
Fucus-levissä ei ole sporofyyttejä. klo Fukusa miehen ja naisen sukuelimet - anteridia siittiöineen ja konseptakkelit, joissa on oogonia ja munat - sijaitsevat talluksen syvennyksissä - scapidit, uros- ja naaraskirvat voivat olla samalla päällä astia tai eri resepteissä. Oogonia istuu häkkijalassa , ilmestyy seinälle käsite(Kuva 32, C, D). Ogoniumsolu jakautuu kolmesti

Riisi. 32. Jäljentäminen Fukusa(mukaan: B. Fott, 1971; W. Braune et ai., 1974): A – talluksen pala (1 – ilmakupla, 2 - resepti); B– viilto astioiden läpi; SISÄÄN– osa naaraspuolisesta scafidiasta; G- oogonia; D- munien vapauttaminen; E– uroskirkkarin leikkaus; JA– haara anteridia; 3 – siittiöiden tuotanto
ytimiä, joista ensimmäinen on pelkistys, jonka seurauksena oogoniumiin muodostuu 8 munaa.
Antheridia muodostuu yleensä haarautuneisiin filamentteihin, jotka tukevat niitä - parafyysejä(Kuva 32, E,JA). Useiden jakautumisten seurauksena muodostuu 64 siittiötä. siittiöt Fukusa silmä sijaitsee pelkistyneen kloroplastin sisällä. Siittiösolun etupäähän muodostuu 13 mikrotubulusta kulkurinykyinen- erityinen rakenne, jonka oletetaan osallistuvan naispuolisen seksuaalisen houkuttimen määrittelyyn.
Siittiöt houkuttelevat munaa Fukusa fucoserraten, munan erittämä epäspesifinen feromoni. Lajitason tunnistaminen liittyy tiettyihin oligosakkarideihin munassa ja siittiössä. Munan pinnalla on erilaisia alueita, joista jokainen sisältää erityisiä orgaanisia yhdisteitä - glykoproteiineja. Siittiöt sisältävät glykoproteiineja, jotka on organisoitunut paikkoihin etusiiman kalvolla ja kasvaimilla sekä itse siittiön pinnalla. Kohdatessaan munasolun siittiö liikkuu munankuoren yli, ikään kuin tutkii sitä etummaisen siimansa avulla. Jos siittiön glykoproteiinit täydentävät munasolun glykoproteiineja, vain munasolu tunnistaa siittiön.
Systematiikka
Keski-Silurialta (noin 425 Ma) tunnetaan fossiilisia löytöjä, jotka voivat liittyä ruskealevään. Mutta nämä löydöt muistuttavat myös joitain nykyajan viher- ja punaleviä. Fossiililöydöt, jotka voidaan ehdottomasti yhdistää nykyaikaisiin ruskealevisiin, ovat peräisin mioseenista (5–25 miljoonaa vuotta). Nämä levät muistuttaa nykyaikaista Dictiotaa. Molekyylimenetelmin määritetty ruskealevien ikä on 155–200 miljoonaa vuotta.
Ruskealeväluokkien erottamiseen käytetään talluksen organisaatiotyyppiä, pyrenoidin läsnäoloa tai puuttumista, kasvutapaa, sukupuolisen lisääntymisen tyyppiä (isogamia, heterogamia, oogamia) ja elinkaarta. Viime vuosina ruskealeväjärjestelmää on tarkistettu aktiivisesti. Eri luokittelujärjestelmissä ruskealevän luokan osana erotetaan 7-14 luokkaa.
Desmarestilainen järjestys–Desmarestiales. Levät ovat tuuheaisia, haaroittuneita, ja niissä on terete, litteä tai litteä tall. Kiinnittyy alustaan hyvin kehittyneellä pohjalla. Talluksen pinnalle kehittyy karvoja.
Suku Desmarestia (kuva 33). Ruskeat tai oliivipensaat, joilla on hyvin erottuva pääverso, jopa 0,8–1 m korkea. Kiinnitetty kartiomaisella pohjalla. Haaroittuvat 3-4 tilausta, vuorotellen tai vastakkain. Viimeisten luokkien oksat ovat litteitä tai harvemmin lankamuotoisia. Lajit ovat merellisiä, elävät arktisilla ja lauhkeilla leveysasteilla 1–20 metrin syvyydessä, muodostaen pensaikkoja tai kasvavat yksittäin.

Riisi. 33. Desmarestia (r. Desmarestia)
Ralphsian järjestys–Ralfsiales. Järjestön edustajilla on tallus kuorien muodossa erilainen rakenne tai koostuvat tyvikuoresta ja pystysuorasta versosta.
Suku Analipus. Tallus koostuu monivuotisesta kuorimaisesta tyviosasta ja siitä ulottuvista yksivuotisista, oliivinruskeista, korkeintaan 40 cm korkeista pystysuoraista versoista Monisoluiset itiöt ovat muodoltaan lieriömäisiä ja rakenteeltaan kaksiriviä. Yksisilmäiset itiöt ovat soikeita ja kehittyvät assimilaatiohaarojen joukossa. Suvun lajit ovat laajalle levinneitä lauhkeiden leveysasteiden merillä muodostaen vöitä vuorovesivyöhykkeellä.
Tilaa Dictyota–Dictyotales.Laukun lajeille on ominaista apikaalinen kasvu. Vegetatiiviset solut sisältävät lukuisia kiekon muotoisia kloroplasteja ilman pyrenoideja. Parenkymaalinen tallus, isomorfinen sukupolvien vuorottelu on ominaista. Sukupuoliprosessi on oogaminen, suvuton lisääntyminen liikkumattomilla itiöillä, siittiöissä on vain yksi siima. Siima on pitkä, siipikarvainen ja eteenpäin suunnattu. Luokan edustajia löytyy pääasiassa trooppisista ja harvemmin lauhkeista meristä.
Suku Dictyota (liite, 4A). Sillä on litteä, oliivinruskea, kaksijakoisesti haarautunut, jopa 11–15 cm korkuinen pensas, jonka oksat ovat suoraviivaisia, kapeita, suonittomia, latvahaarukkaisia. Tetrasporangiat muodostavat talluksen pinnalle soreja, joilla on epämääräinen tai rengasmainen muoto. Levitetty lauhkeiden ja lämpimien leveysasteiden merivesissä.
Tilaa Laminaria–Laminariales. Laminarialla on tallus, joka koostuu levystä, varresta ja kiinnityselimistä - pohjasta tai risoideista. Levy on tasainen tai ryppyinen, kokonainen tai leikattu, reikiä, poimuja, petsejä tai ilman niitä, useista kymmenistä senttimetreistä 20–40 m pitkä Varsi on haarautunut tai useammin haarautumaton. Sporofyytit ovat yksivuotisia tai monivuotisia, jotkut lajit ovat 11–18-vuotiaita. Limakanavat sijaitsevat monien rakkolevän kuoressa.
Ruskealevien suosituimmat edustajat ovat rakkolevä (sukutyypit Laminaria tai Saccharina), tai "merilevää", joita levitetään pääasiassa Tyyni valtameri. Niitä löytyy veden pinnasta (reunasta) 200 m syvyyteen asti. Laminaria kasvaa mieluiten 2–20 (30) m syvyydessä, paikoissa, joissa vesi liikkuu jatkuvasti, kiinnittyen kiviin ja kiviin. Laminariat muodostavat levävyöhykkeitä Kaukoidän merien rannikolla. Voimakkaimmat pensaat löytyvät Kurilien saarilta ja noin klo. Hokkaido, jonne noin puolet maailman laminariavaroista on keskittynyt. Maailman rakkoleväsato saavuttaa 2 miljoonaa tonnia tuorepainoa, kun taas yli miljoona tonnia tulee sen meriviljelyn tuotannosta Kiinassa.
Suku Sakariini (Laminaria). Tallus on 0,5–5–10 m pitkä, siinä on paksu kiiltävä levy, elastinen varsi ja tiiviisti toisiinsa kietouttuneet risoidit, jotka on kiinnitetty erilaisiin kiinteisiin alustoihin. Levyn reunat ovat sileät tai aaltoilevat, levyn pinnassa on usein pullistumia ja taitoksia. Levyjen molemmille puolille muodostuu itiöpesäkkeitä - sori -, itiöiden vapautumisen jälkeen levyn yläosa tuhoutuu. Elää kahdesta useaan vuoteen. Sakariinit ovat arvokas teollisuuden raaka-aine ja meriviljelyn kohde. Saccharina japanilainen muodostaa pensaikkoja Japaninmeren pohjoisosassa, samoin kuin lähellä Sahalinin etelä- ja kaakkoisrannikkoa ja Etelä-Kuriilisaaria. Saccharina japanilainen on arvokkain kaupallinen laji, sillä on erinomainen maku ja sitä käytetään erilaisten ruokien, säilykkeiden ja makeisten valmistukseen.
Tilaa Sphacelariae–Sphacelariales.
Tähän lahkoon kuuluvilla ruskeilla levillä on haarautuva talli, jossa on apikaalista kasvua. Karvaiset oksat. Vegetatiivisissa soluissa on lukuisia kloroplasteja ilman pyrenoideja. Enemmistön elinkaaren aikana isomorfinen sukupolvien vaihto, seksuaalinen prosessi on isogamia tai heterogamia. Levät muodostavat pieniä, enintään 3 cm korkeita ruskeita rypäleterttuja ja jopa 20 cm korkeita haarautuneita pensaita kiville tai muille leville; on endofyyttisiä edustajia.
Suku Sphacelaria. Sillä on ohut tallus. Haaroittuvat filamentit muodostavat pieniä, jopa 0,5–3 cm korkeita, ruskeita kimppuja. Vegetatiivisen lisääntymisen suorittavat erikoistuneet, usein kolmipalkiset sikiön oksat - leviävät. Seksuaalinen prosessi on isogaminen. Naisten sukusolut erittävät feromoneja ectocarpen ja multifiden. Suvun lajeja on sekä polaarisilla että trooppisilla leveysasteilla.
Tilaa Scytosiphon–Scytosiphonales. Edustajat, joilla on suolisto-, rakkulaarinen, lamellimainen tai sakkulaarinen, usein ontto talli, jonka pituus on enintään 25 cm. Kuori on yksirivinen tai kaksirivinen, talluksen sisäosa (seinä) muodostuu useista kerroksista suuria soluja. Talluksen pinnalle kehittyy monisoluisia itiöitä, jotka kerääntyvät soriin, jotka tukevat niiden lankoja - parafyysejä ja hiuskimppuja.
Suku Scytosiphon on ohutseinäinen ruskea 4–25 cm pitkä putki (Liite 4B). Tallus on kiinnitetty pienellä pohjalla ja siinä on harvinaisia poikittaisia supistuksia. Sitä esiintyy merien rannikkovyöhykkeellä eri leveysasteilla, muodostaen usein vyöhykkeitä vuorovesivyöhykkeellä.
Fucus tilaus–Fucales. Heillä on monivuotinen talli suurten, epäsäännöllisesti tai kaksijakoisesti haarautuvien, jopa 1–3 metrin pituisten pensaiden muodossa. Ne on kiinnitetty alustaan kartiomaisella pohjalla. Talluksessa on versoja, rypäleitä (lehtimäisiä muodostelmia), säiliöitä (sukusolujen säiliöitä) ja uivia (ilma) rakkoja. Niitä löytyy kaikkien lauhkean ja arktisten merien vuorovesivyöhykkeeltä, missä ne muodostavat voimakkaita paksuja - vöitä.
Suku sargassum(Kuva 34, SISÄÄN). Runsaan haarautuneet oliivinruskeat tai ruskeat pensaat, korkeus 0,5–1,5 m, kiinnitetty kartiomaisella pohjalla. Fylloidit kehittyvät sivuversoissa, jotka ulottuvat lieriömäisestä varresta talluksen ala- ja keskiosissa. Phylloidit ovat nahkamaisia, keskikylkeä, 0,2–2 cm pitkiä. Uimarakot ovat soikeita tai pyöristettyjä, kasvuston kanssa tai ilman. Sargassumit ovat laajalle levinneitä lauhkeilla ja subtrooppisilla leveysasteilla alemmasta rannikosta 10 metrin syvyyteen, missä ne muodostavat tiheitä tai harvoja metsikköjä. Eteläisellä Tyynellämerellä - niin kutsutulla Sargassomerellä - ne muodostavat valtavia kelluvia kasaumia, joissa ne lisääntyvät vain kasvullisesti.
Suku Fucus (kuva 32).Thallus nahkainen, tuuhea, oksat kaksijakoisesti, 25–50 cm korkeat.Lähellä Kaukoidän merten rannikkoa, kasvaa useita vuosia, elää rantavyöhykkeellä. Muodostaa usein suuria paksuja matalassa vedessä. Tämän suvun lajeille on ominaista kaksijakoisuus
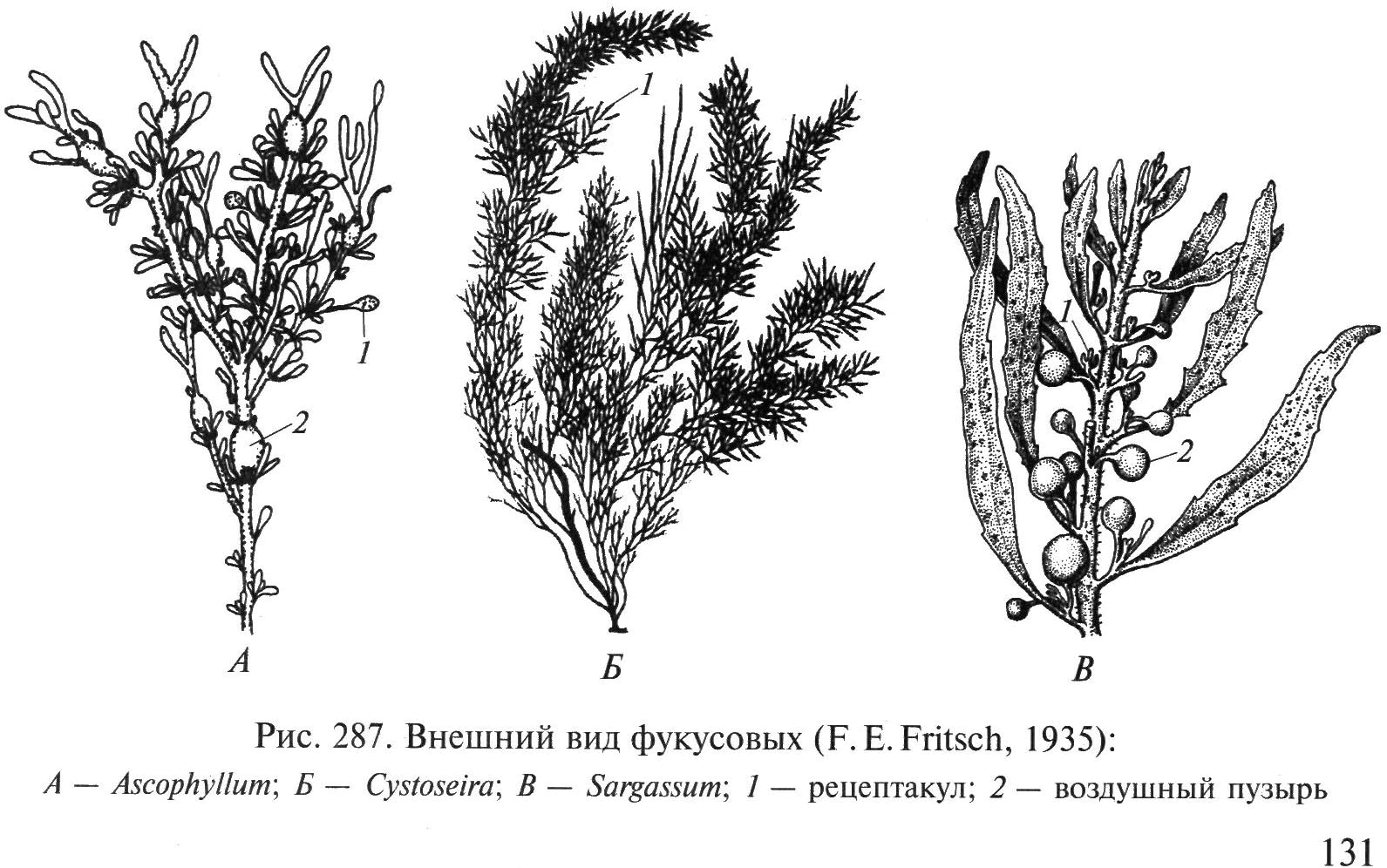
Riisi. 34. Fucus-levien ulkonäkö: A – Ascophyllum; B – Cystoseira; SISÄÄN – sargassum; 1 - astia, 2 - uimarakko
haarautunut tallus litteillä oksilla, jossa on pitkittäinen kylkiluo, joka on kiinnitetty kiviin kartiomaisella pohjalla. Fucuseja käytetään lannoitteina, karjan rehuna, rehujauhon, alginaattien valmistukseen, käytetään lääketieteessä struuman hoitoon, painonkorjaukseen jne.
Tilaa Ectocarps–Ectocarpales. Tähän sisältyvät levät ovat mikroskooppisia, vapaasti eläviä, epifyyttejä, epitsoiitteja tai endofyyttejä. Sukuelimiä edustavat yksisilmäiset (sporangiat) ja monisilmäiset (gametangia) itiöt; ne ovat pitkiä lieriömäisiä, lyhytsylinterimäisiä tai tyynyn muotoisia.
Suku ectocarpus. Tallus pehmeiden ruskeiden pensaiden muodossa, korkeintaan 2 cm. Holkit muodostuvat yksirivisistä, vuorotellen, kaksijakoisista tai väärin haarautuneista lankoista. Monisoluiset itiöt ovat palomaisia, muodostuneet sivuhaarojen kylkeen 1–3-soluiseen varteen. Lajeja esiintyy eri leveysasteilla olevissa merissä, puhtaissa ja saastuneissa vesissä, rannikolla ja sublitoraalin yläosassa. Epifyyttejä löytyy myös ihmisperäisten substraattien likaantumisesta.
Ekologia
Suurin osa ruskealevistä elää merivesissä. Vain 8 lajia tavataan makeissa vesissä. Siellä on sekä yksivuotisia että monivuotisia lajeja, joiden ikä voi olla 15–18 vuotta. Laminaria levä Nereocystis Ja Makrocystis, jotka elävät Pohjois-Amerikan Tyynenmeren rannikolla, ovat kaikkien levien enimmäiskoko - jopa 30–40 m pitkiä. Ne muodostavat jättimäisiä vedenalaisia metsiä Kaukoidän merissä. Ruskeat levät kiinnittyvät monenlaisiin substraatteihin - kiviin, kiviin, soraan, merieläinten kuoriin ja kuoriin sekä muihin leviin. Jotkut ruskealevän pienet muodot elävät muiden levien kudoksissa endofyytteinä. Useimmat ruskeat lajit elävät kiinnittyneessä tilassa. Maasta irti repeytyneet tallit kulkeutuvat virran mukana tyynille mutapohjaisille paikoille ja ovat siellä edelleen olemassa. Lajit, joilla on ilmakuplia talluksessa, irtautuvat maasta, kelluvat pintaan muodostaen suuria kelluvia klustereita erityisesti alueilla, joilla on vakaa ympyrävirtaus, kuten Sargassomerellä. Ne esiintyvät tällaisissa klustereissa melko pitkään ja lisääntyvät vain kasvullisesti.
Ruskealevät ovat laajalle levinneitä kaikissa merissä, mutta ne saavuttavat suurimman kehittymisensä lauhkeiden ja alipolaaristen leveysasteilla. Niiden suuret tallit kasvavat kiinnittyneenä kallioihin ja kiviin, ja hiljaisissa paikoissa lähellä rannikkoa ja suurissa syvyyksissä niitä voidaan pitää jopa nilviäisten kuorien ja soran venttiileissä. Voit tavata niitä eri syvyyksillä - rantavyöhykkeeltä, jossa laskuveden aikaan ne ovat poissa vedestä tuntikausia, jopa 40–200 m. Havaijin saarten alueella 180 metrin syvyydessä lajit suvusta sargassum, A Rakkolevä V Adrianmeri löytyi 200 m:n syvyydestä. Runsaat ruskealeväpeikot havaitaan 6–15 m:n syvyydessä, missä ovat parhaat valaistusolosuhteet ja jatkuva veden liike, joka tuo ravinteita niiden talliin ja rajoihin. kasvissyöjien asuttaminen - fytofagit.
Luokka kultalevät -Chrysophyceae
Yksisoluiset, siirtomaa- tai monisoluiset organismit, joiden elinkaaressa on yleensä siimavaihe. Solut ovat yksitumaisia, ja niissä on yksi tai useampi kloroplasti. Stigmaa esiintyy yleensä (kuva 36). Flagella 1–2, eriarvoinen. Supistumisvakuoli sijaitsee solun etuosassa. Näiden organismien solujen ulkokuoret ovat hyvin erilaisia. Primitiivisimmissä lajeissa solu on pukeutunut herkälle periplastille, joka mahdollistaa erimuotoisten ulkonemien muodostumisen (rhizopodia, pseudopodia), toisissa plasmalemma on peitetty kovalla selluloosakalvolla, joskus voimakkaasti limaisella. Kultalevissä olevia taloja on eri muotoisia: maljakon muotoisia, pallomaisia, munamaisia, lieriömäisiä; yhdellä tai useammalla reiällä. Häkki kiinnitetään talon pohjaan joustavalla jalalla tai voi olla vapaa. Joissakin krysofyyteissä solun pinnalle kertyy erimuotoisia ja -kokoisia kalkkipitoisia muodostelmia - kokkoliitteja, ja piisiipillä on sisäinen piirunko. Näiden luurankojen rakenteen monimuotoisuus mahdollistaa fossiilisten piiflagellaattien eri muotojen jäänteiden käyttämisen niitä sisältävien geologisten kivien iän määrittämiseen. Kehittyneimmillä kultaleväosaston edustajilla solut on puettu kuoreen, joka koostuu piisuomuista (joskus on piikkejä) tai suljettu taloihin, joiden aukkojen kautta tulee ulos lippuja tai pseudopodia.
Kultalevälajeihin kuuluvat talot ovat maljakon muotoisia, pallomaisia, munamaisia, lieriömäisiä, ja niissä on yksi tai useampi reikä. Häkki kiinnitetään talon pohjaan joustavalla jalalla tai voi olla vapaa.
Pigmentit - klorofyllit A Ja Kanssa, fukoksantiini. Krysofyyteille on ominaista korkein fukoksantiinipitoisuus heterokonteista (55-93 % kaikista karotenoideista).
Suurin osa lajeista on makean veden fototrofeja, mutta myös heterotrofeja ja fagotrofeja löytyy.
Vara-aineet - krysolaminariini ja öljyt.
Kultalevien yksisoluisista muodoista vallitsevat makean veden planktonlajit ja monisoluisista pohja- tai epifyyttiset lajit. Niitä löytyy pääasiassa viileistä merestä ja makeista vesistöistä. Noin 360 lajia tunnetaan.

Riisi. 36. Kultalevät (C. Hoek van den et al., 1995 mukaan): A- solun rakenne; B- radikulaarinen järjestelmä; SISÄÄN- kolmiosainen mastigonema; G- kloroplastin osan leikkaus; 1 - perusrunko; 2 - tyviturvotus; 3 - lyhyt siima; 4 - stigma; 5 - risoplasti; 6 - ydin; 7 - ydinkalvo; 8 - ydin; 9 - mitokondrio; 10 - vakuolit; 11 - lima; 12 - plasmalemma; 13 - rakkulat; 14 - limakalvo; 15 - lipidit; 16 - kloroplasti; 17 - Golgi-laitteet; 18 - supistuva vakuoli; 19 - pitkä flagellum: 20 - 1. juuri; 21 - 2. selkäranka; 22 - 3. selkäranka; 23 - 4. juuri; 24 - kloroplastinukleoidi; 25 - HES; 26 - kloroplastikuori; 27 - vyön lamelli; 28 - lamelli; 29 - pitkä lateraalinen filamentti; 30 - lyhyt lateraalinen filamentti; 31 - päätelanka; 32 - mastigoneeman putkimainen osa
talluksen rakenne kultalevissä se on useimmiten monadinen, mutta voi olla hyvin monimuotoinen: kokkoidinen, palmelloidinen, plasmoidinen, ameboidinen, rihmamainen, parenkymaalinen (kuva 37).

Riisi. 37. Kultalevät (L.L. Velikanov et al., 1981 mukaan): A – Ochromonas: 1 - ulkonäkö, 2 - kysta; B – Chromulina: 1 - ulkonäkö, 2 - kysta, 3 - kaavio kalvon muodostumisesta kystasta; SISÄÄN – Dinobrion: 1 - yleinen näkymä pesäkkeestä, 2 - kysta; G – Chrysameba; D – gibberdia; E – Hydrurus: 1 - yleiskuva pesäkkeestä, 2 - haaran kärki, 3 - zoospore
jäljentäminen. Kultalevissä on kolme lisääntymismenetelmää: vegetatiivinen, aseksuaalinen ja seksuaalinen.
Vegetatiivinen lisääntyminen tapahtuu pitkittäisen solunjakautumisen tai pesäkkeen hajoamisen osiin seurauksena solun jakautumisesta yhteen, kahteen tai kolmeen suuntaan.
seksuaalinen prosessi(kultalevien solut ovat diploideja) - isogamia tai konjugaatio. Seksuaalisen prosessin seurauksena sekä epäsuotuisissa olosuhteissa muodostuu kystat, joissa on paksu piitä sisältävä kuori.
suvuton lisääntyminen yksi- tai kahdella leimautuneella zoosporilla, jotka kehittyvät tavallisissa vegetatiivisissa soluissa tai zoosporangioissa. Seksuaalinen prosessi: holo-, iso- ja autogamia. Hologamia on eräänlainen seksuaalinen prosessi, jossa kaksi vegetatiivisten solujen kaltaista solua sulautuvat etupäistä kaksitumaiseksi tsygootiksi, joka sitten muuttuu silikoituneeksi kystaksi.
Systematiikka
Kultalevistä eristettiin viime vuosisadan lopulla molekyyligeneettisten tutkimusten tulosten perusteella luokka Sinuraceae. Luokka Feotamniaceae erotetaan myös - keltavihreistä ja kultalevistä. Siksi Chrysophyceae-luokan soveltamisalaa on supistettu ja se sisältää tällä hetkellä noin 360 lajia, jotka on luokiteltu kolmeen luokkaan: Chromulinales, Hibberdiales ja Hydrurales. Siipien muoto, rakenne ja lukumäärä sekä pigmentin koostumus ja rungon rakenne ovat tärkeitä systemaattisia hahmoja.
Kromuliinijärjestys - Chromulinales. Tähän luokkaan kuuluvat organismit, joilla on monadinen, palmelloidinen ja ameboidityyppinen talluksen erilaistuminen. Monadiset solut, joissa yksi flagellum näkyy valomikroskoopilla.
Suku Chrysameba(Kuva 37, A) - makeanveden ameboidilevät, niiden elinkaaressa on siimavaihe. Tsygoottista kehittyy stomatokysta.
Suku Chromulina(Kuva 37, SISÄÄN) - yksisoluiset vapaasti uivat organismit, jotka elävät pääasiassa makeissa vesissä. Solut ovat alastomia; on yksi tai kaksi kultaista plastidia.
Suku Dinobrion(Kuva 37 , G)- yksisoluiset ja siirtomaa-, vapaasti kelluvat tai kiinnittyneet edustajat. Monadit sijaitsevat maljakkomaisissa taloissa, jotka muodostuvat selluloosamikrofibrillistä. Selluloosan lisäksi talossa on myös suuri määrä aminohappoja. Pesäkkeiden tuuhea muoto liittyy vegetatiivisen solunjakautumisen menetelmään, kun yksi vanhempainkodista lähtevistä tytärsoluista kiinnittyy sen reikään. Pyöriessään akselinsa ympäri se muodostaa oman talonsa. Molemmat lapsisolut voivat myös poistua vanhemmista. Solut sisältävät 1-2 kloroplastia, silmä ja 2 supistuvaa vakuolia näkyvät solun etupäässä. Krysolaminariini sijaitsee tyhjiössä solun takapäässä. Lisääntyminen on vegetatiivista ja seksuaalista.
Tilaa Gibberdiae–Hibberdiales. Tämän luokan edustajille on ominaista ainutlaatuinen pigmenttikoostumus. Fukoksantiinin lisäksi niissä on lisäksi valoa keräävä karotenoidipigmentti, anteraksantiini.
Sisältää synnytyksen Gibberdia, Chromophyton, Styloceras, Chrysopixis, Platyteka ja muut.
Suku gibberdia sen elinkaaressa on kaksi vaihetta: siirtomaa-palmelloidi liikkumaton ja yksisoluinen monadinen liikkuvuus. Valomikroskopialla vain yksi flagellum on näkyvissä monadisessa vaiheessa. Fukoksantiinin lisäksi on ominaista toisen lisävaloa keräävän karotenoidipigmentin, anteraksantiinin, läsnäolo.
Tilaa Hydrorus–Hydrurales. Tämän luokan edustajilla on tallus, jonka rakenne on palmelloidinen ja pseudoparenkymaalinen. Ainutlaatuisten tetraedristen zoosporien esiintyminen on ominaista. Ei ole silmää. Mitoosi on puolisuljettu.
Sisältää synnytyksen Hydrorus, Celloniella, Federmatium, Chrysonebula ja muut.
Suku Hydrurus(Kuva 37, E) on palmelloidilevistä erottuvin. Sen tallus on suuria, jopa 30 cm pitkiä limaisia pesäkkeitä, jotka näyttävät ruskeilta haarautuneilta naruilta, jotka usein lähettävät erittäin epämiellyttävää hajua. Pesäkkeissä voidaan erottaa päärunko ja sivuhaarat. Tavalliseen limaan upotetut solut sisältävät yhden kupinmuotoisen kromatoforin ja useita supistumisvakuoleja. Solut sijaitsevat löyhästi pesäkkeen reunalla ja tiheämpiä keskellä. Tallus voi kasvaa vain jakamalla apikaalisia soluja. Aseksuaalisen lisääntymisen aikana zoosporeja muodostuu pesäkkeen sivuhaarojen soluihin. Muodostaa pallomaisia kystoja. Sitä esiintyy vuoristopuroissa ja joissa, joissa on kylmää vettä, missä se kiinnittyy koviin alustoihin.
Luokka Sinura-levä -Sinurophyceae
syn- yhdessä ja ura- häntä. Tämä organismiryhmä eristettiin vuonna 1987 kultalevistä. Se yhdistää monadisia yksittäisiä ja siirtomaaorganismeja, toisinaan vuorotellen monadisten ja palmelloidisten vaiheiden elinkaaressa. Pektiinikuorten pinta on peitetty piihiutalekuorella. Mitokondriot, joissa on putkimaisia cristae, sijaitsevat yleensä sytoplasmassa kloroplastin ympärillä. Ydin on yksi, kloroplasteja, joita ympäröi neljä kalvoa, yleensä kaksi. Lamellit ovat trityylakoidisia, niissä on vyölamelli. Pääpigmentit ovat klorofyllit A Ja Kanssa, β -karoteeni ja fukoksantiini. Ei ole stigmaa. Solut, joissa on yleensä 2 erilaista siimoa. Pitkä siima on suunnattu eteenpäin. Lyhyt, sileä siima, joskus voimakkaasti kaventunut, suuntautunut taaksepäin.
Solut lisääntyvät pääasiassa pitkittäisjakautumalla. Hajotessaan yhdyskunnat synnyttävät nuoria pesäkkeitä. Joissakin lajeissa seksuaalista prosessia kuvataan isogamiana. Samanaikaisesti, toisin kuin kultalevässä, isogamettien fuusio sinilevissä ei tapahdu etu-, vaan niiden takapäissä. Silicified kystat, joissa on huokoset, muodostuvat endogeenisesti, samanlaisia kuin krysofyyttien.
Pääasiassa planktoniset muodot, palmelloidivaiheet ovat osa pohjaeliöstöä. Useimmat sinuralevälajit ovat fototrofisia makean veden organismeja.
Systematiikka.
Sinura-leväluokkaan kuuluu 7 luokkaa: Chloramoebales, Synurales, Rhizochloridales, Ochromonadales, Heterogloeales, Parmales ja Thallochrysidales.
Tilaa Sinura - Synurales. Monadiset muodot, pektiinisolukalvot on yleensä peitetty piidioksidihiutalekuorella, sementoituna orgaanisella aineella yhdeksi koteloksi. Kloroplastit yksi tai kaksi.
Suku Sinura(Kuva 38) - makeanveden monadiset muodot, joissa on kaksi epätasa-arvoista flagellaa ja kaksi parietaalista kloroplastia. Ydin on päärynän muotoinen, sijaitsee solun etuosassa. Ytimen takana on yksi suuri tyhjiö ja solun takaosassa useita pieniä supistumisvakuoleja. Solut ovat suomujen peitossa, kuten laatat, lisääntyminen on useimmiten vegetatiivista, joidenkin sukupuoliprosessi tunnetaan. Muodostaa pesäkkeitä (kuva 39, A). Usein kystat näkyvät kaikissa pesäkkeen soluissa. Massakehityksellä Sinura voi aiheuttaa veteen epämiellyttävän hajun.

Riisi. 38. Sinura-solujen ulkonäkö
Tilaa Rhizochloride - Rhizochloridales. Pääasiassa makean veden organismeja, joilla on rhizopodiaalinen talluksen erilaistuminen ja jotka elävät pääasiassa makeassa vesistössä.
Suku Mallomonas(Kuva 39, B-D) – yksisoluinen monadi, jossa on yksi hyvin merkitty siima ja yksi kaksihaarainen kloroplasti.

Riisi. 39. Sinuurilevien ulkonäkö (G.A. Belyakova et al., 2006 mukaan): A – halkeavaa siirtokuntaa Sinura; B, C – statosporien muodostuminen ja G – ulkomuoto Mallomonas
Solu on peitetty kaakeloiduilla ja kierteisillä suomuilla, joista osassa on neulamaisia piikkejä (kuva 39, G). Statosporien esiintyminen on ominaista (kuva 39, B, C). Useille edustajille kuvataan seksuaalista prosessia, hologamiaa.
Suku Mixochloris elää sphagnum sammalten lehtien soluissa, näyttää suurelta moniytimiseltä plasmodiumilta. Se muodostaa syksyllä kystat, jotka itävät keväällä. Niistä nousevat eläinporit tai amebat, jotka tunkeutuvat tyhjiin lehtisoluihin ja sulautuvat siellä plasmodiumiksi.
Tilaa Chloramoebaceae - Chloramoebales. Sisältää monadin edustajat. Löytyy suolaisista ja makeista vesistä.
Suku heterochloris – sen solut voivat muuttaa muotoaan muodostaen pseudopodia. Tämä ilmiö on luontainen monille järjestyksen edustajille, samoin kuin taipumukselle eläinravitsemustapaan. Solussa on useita kloroplasteja, öljypisaroita ja krysolaminariinia. Solun edessä on supistuvia vakuoleja, keskellä - yksi ydin. Lisääntyy vegetatiivisesti solujakautumalla.
Tilaa Ochromonadales - Ochromonadales. Yhdistää paljaat muodot kahteen epätasa-arvoiseen siimaan. Makean veden ja meren muodot.
Suku Ochromonas sisältää yksisoluisia monadeja, joissa on kaksi eriarvoista siimat. Solut peitetään vain plasmalemmalla. Solun etupäässä on supistuva vakuoli ja silmä, takapäässä krysolaminariininen tyhjiö. Se lisääntyy vegetatiivisesti, fissioura alkaa solun etupäästä kahden flagellaarisen emäsparin välistä. Suvun lajit ovat yleisiä oligotrofisissa makeissa vesissä, mutta siellä on myös meren edustajia.
Tilaa Heterogleyes - Heterogloeales. Sisältää levät, joilla on palmelloidityyppinen talluksen erilaistuminen. Luokan edustajia tavataan useammin makeissa kuin suolaisissa vesissä.
Suku Helmintoglea asuu murtovesissä. Tämä laajennetulla pohjalla istuva siirtokunta koostuu haarautuvista limaisista nauhoista. Protoplastit ovat satunnaisesti asettuneet tähän limaan, joista jokaista ympäröi oma limakalvo.
Phaeotamnia leväluokka - Phaeothamniophyceae
Luokan nimi tulee tyyppisuvusta Phaeothamnion(kreikasta. phaeos- tumma pensas). Tämän organismiryhmän edustajat tunnistettiin kelta-vihreistä ja kultalevistä erillisessä luokassa vuonna 1998 ribosomaalisten geenien sekvenssin ja solun ultrarakenteen ominaisuuksien analyysin perusteella. Feotamnialevälle on ominaista ainutlaatuinen pigmenttien yhdistelmä: fukoksantiini heteroksantiinin kanssa, violaksantiinin puuttuminen.
Solurakenteen ominaisuudet: vakuolien puuttuminen krysolaminariinilla ja endogeenisten kystien puuttuminen piidioksidiseinämillä.
Luokkaan kuuluvat yksisoluiset, siirtomaa- ja monisoluiset organismit kokkoidinen, palmelloidinen ja rihmamainen tallusrakenne. Leimautuneet vaiheet, joissa on kaksi eriarvoista siimoa. Flagella lateraalinen tai subapical. Stigmaa löytyy eläinporeista. Pigmentit - klorofyllit A Ja Kanssa, β -karoteeni, fukoksantiini, diadinoksantiini, diatoksantiini ja heteroksantiini. Päävaratuote on paramyloni ( β -glukaani). Soluseinä on läsnä, solunjakautumisen aikana tytärseinä muodostuu emoseinän sisään.
Feotamniaceae-lajeissa tärkeimmistä lisääntymismenetelmistä löytyy vain vegetatiivisia ja aseksuaalisia. Aseksuaalinen lisääntyminen tapahtuu autoitiöiden tai zoosporien avulla. Sukupuolista lisääntymistä ei tunneta.
Feotamnia-levälajit elävät yksinomaan makeissa vesissä. Systematiikka.
Tällä hetkellä feotamnialevien monofyleettinen luokka on vahvistettu, ja okrofyyttien joukossa se on lähempänä ruskea- ja keltaviherleviä kuin kultaleviä. Tällä hetkellä näistä levistä tunnetaan noin 30 lajia, jotka johtuvat yhdestä lahko Theotamnia - Phaeothamniales.
Suku Theotamnion edustaa kiinnitettyjä haarautuvia lankoja, joiden korkeus on enintään 1 cm (kuva 40). Solut ovat sylinterimäisiä, epäsäännöllisiä, ylöspäin laajentuneita

Riisi. 40. Ulkonäkö Theotamnion.
kaksi tai kolme kertaa leveää pidempi. Solunjakautumisen aikana emoseinämän materiaalia kuluu filamenttisoluja ympäröivän kerrostetun vaipan muodostumiseen. Solun reunalla plasmakalvon alla on rakkuloita. Ne muistuttavat fysodeja - muodostumia, joita löytyy ruskealevien soluista. Solut sisältävät yhdestä useisiin kiekon muotoisia oliivinruskeita plastideja. Zoosporeja muodostaa 1-2, harvemmin 4-8 yhdessä solussa. Zoosporeissa flagellat ovat kiinnittyneet sivusuunnassa. Siimajuurten sijainti Theotamnion muistuttaa kelta-vihreitä ja ruskealeviä. Siitä puuttuu myös rhizoplast. Elinkaaren aikana kystat voivat muodostua, mutta niiden seinissä ei ole piitä. Kystat itävät zoosporeina. Laskeutuu rihmamaisten levien epifyyttinä. Asuu pysähtyneissä ja hitaasti virtaavissa makeissa vesistöissä.
Luokka Rafid levät -Raphidophyceae
Luokan nimi tulee kreikan kielestä rafidi - neula. Yhdistää yksisoluisia kaksisoluisia organismeja, joilla ei ole soluseinää. Enimmäkseen makean veden organismeja, joiden solut ovat litistyneet (kuva 41).
Soluilla on eripituisia siimoja, eteenpäin suunnattu siima on pinnallinen ja pidempi, taaksepäin suunnattu siima on sileä ja lyhyempi. Siirtymäkierre puuttuu. Kloroplastit ovat pieniä, lukuisia tai harvemmin 1-2 per solu, ja niitä ympäröi neljä kalvoa, joista kaksi on CES-kalvoja. Tylakoidit on järjestetty kolmen pinoihin. Ydin on suuri, ja sitä ympäröi diktyosomirengas. Mitokondriot putkimaisilla risteillä. Siellä on supistuvia vakuoleja. Sytoplasma on usein vakuoloitunut. Solut ovat alastomia, ja niitä ympäröi vain plasmalemma. Stigma (silmä) puuttuu yleensä.
Fotosynteettiset pigmentit - klorofyllit A Ja Kanssa, β -karoteeni, vosheriaksantiini, dinoksantiini, diadinoksantiini, heteroksantiini.
Vara-assimilaatiotuotteita ovat rasvat ja öljyt, harvemmin tärkkelys ja krysolaminariini.
Ne elävät myös sfagnum-soissa ja muissa elinympäristöissä, joissa on happamia ja neutraaleja vesireaktioita. Jotkut lajit löytyvät murto- ja merivesistä.

Riisi. 41. Rapidlevien ulkonäkö.
Systematiikka.
Hyvin pitkään tätä leväryhmää pidettiin sen vihreän värin, nielun läsnäolon ja useiden muiden rakenteellisten ominaisuuksien vuoksi euglenoidien joukossa Chloromonas-luokkana. Mutta sytologian, biokemian, fysiologian ja molekyylibiologian tiedot osoittivat niiden kiistattoman kuulumisen Heterocont (Ochrophyte) leväjakoon ja Rafid-leväryhmän monofyleettisen luonteen.
Ainoastaan tunnetaan noin 25 lajia tilaa Huttonellaceae– Chattonellales.
Suku heterosigma(Kuva 42, A) sisältää merisiimat. Soikeat solut, joissa flagella on siirtynyt kohti sivupuoli, plastidit sijaitsevat solun reunalla. Trikokystiä ei ole. Ne uivat ja tekevät pyöriviä liikkeitä kehon pituusakselin ympäri. Saattaa aiheuttaa myrkyllisiä "vesikukintoja" rannikkovesissä.

Riisi. 42. Rafid levät (tekijä: R. E. Lee, 1999): A – heterosigma; B – Goniostomum; 1 - kloroplasti; 2 - mukokysti; 3 - supistuva vakuoli; 4 - trikokysti; 5 - ydin
Suku Goniostomum(Kuva 42, B) ovat liikkuvia dorsoventraalisesti puristettuja monadeja. Selkäpuoli on kaareva, vatsapuoli litistynyt. Etupäässä oleva uurre johtaa kolmiomaiseen nieluun, josta kaksi pitkää siimat nousevat esiin; niiden pituus on verrattavissa solun pituuteen. Plastidit sijaitsevat solun reunalla. Plasmalemman alla on trikokystoja. Asuu makeissa vesissä, joiden pH on hapan.
Luokka Eustigma levät -Eustigmatophyceae
Luokan nimi tulee kreikan kielestä eu- "hyvä ja stigma- "merkki", "piste". Luokka yhdistää paljaat yksisoluiset, harvemmin siirtomaa-organismit, joilla on pääosin kokkoidirakenne.
Yksi tai useampi ydin. Kloroplasti on yleensä yksi, jota ympäröi 4 kalvoa. Siima on yleensä yksi etuosa ja toinen tyvirunko tai harvemmin kaksi epätasaista siipiä. Stigma (silmä) on läsnä, sijaitsee kloroplastin ulkopuolella, sen rakeet ovat ilman kalvoja.

Riisi. 43. Eustigma-levien ulkonäkö
Kloroplasti on yleensä yksittäinen, suuri, moniliuskainen, kuppimainen tai parietaalinen, väriltään kelta-vihreä. Tylakoidit on ryhmitelty lamelleiksi kolmen ryhmän ryhmiin; ympäröivä lamelli puuttuu.
Pääpigmentit ovat klorofylli a, β-karoteeni, violaksantiini ja vosheriaksantiini. Pyrenoidi esiintyy yleensä vain vegetatiivisissa muodoissa.
Vara-aineet ovat öljyjä ja luonteeltaan tuntemattomia aineita, jotka kerrostuvat kiinteänä aineena plastidien ulkopuolelle. Tärkkelystä ei muodostu.
Ne elävät makeissa vesissä, ovat vähemmän yleisiä merissä ja maaperässä.
Systematiikka.
Aikaisemmin tämä organismiryhmä luokiteltiin keltavihreiksi leviksi. Tällä hetkellä Eustigma-leviä pidetään luokkana Heterocontophytes-osastolla. Noin 35 lajia tunnetaan, enimmäkseen makean veden, katsotaan ainoaksi Eustigma järjestys - Eustigmatales.
Suku Eustigmatos(Kuva 43) - yksisoluiset pienet pallomaiset levät. Soluseinä on sileä, ehjä, ilman koristeita. Keltaisenvihreä kloroplasti yksittäinen, lohko, parietaalinen, varrellisia monitahoisia pyrenoideja. Suuri keskusvakuoli punaisella sisällöllä. Se lisääntyy kahdella tai neljällä autoitiöllä tai pullon muotoisella zoosporilla, joissa on yksi sisääntuleva siima, suuri stigma, joka sijaitsee kloroplastin ulkopuolella, etupäässä ja yksi posteriorinen kloroplasti ilman pyrenoidia. Maaperän kasviston yleinen komponentti. Tunnetaan Uudesta-Seelannista, Itävallasta, Islannista ja Arizonan altailta "sinappimerilevänä".
Luokka Kelta-vihreät levät -Xanthophyceae
Keltavihreät levät ovat leviä, joiden kloroplastit ovat värjätty vaaleaksi tai tummaksi keltainen, hyvin harvoin vihreä ja vain satunnaisesti sininen. Tallin värin määrää seuraavien pigmenttien läsnäolo kloroplastisoluissa - klorofyllit A Ja Kanssa, β -karoteeni ja ksantofyllit. Jälkimmäisen hallitsevuus määrittää kelta-vihreiden levien värin omaperäisyyden. Lisäksi paramylonia, öljypisaroita ja vain joissakin lajeissa lisäksi leukosiinin ja volutiinin kokkareita kerääntyy soluihin assimilaation päätuotteena. Keltavihreät levät eivät muodosta tärkkelystä. Keltaisen vihreän erottuva piirre on monadisen rakenteen läsnäolo vegetatiivisissa soluissa ja kaksi epätasa-arvoista flagellaa zoosporeissa. Soluseinä sisältää selluloosaa, glukoosia ja uronihappoja. Soluseinä koostuu usein kahdesta osasta.
Lisääntyminen on vegetatiivista, aseksuaalista ja seksuaalista.
Levitetty laajalti makeissa vesissä. Harvoin meri-, murtovesi- ja maaperässä.
Aikaisemmin keltavihreäleväluokkaa kutsuttiin Tribophytium leväksi Tribonema-tyyppisuvun mukaan (kreikasta. tribon – taitava, ovela ja nema – lanka). Noin 450 lajia tunnetaan.
Keltaisille vihreille on ominaista merkittävä morfologinen monimuotoisuus. Tämän osaston lukuisten edustajien joukosta löytyy melkein kaikki päätyypit kehon rakenteesta: ameboidi, monadinen, palmelloidi, kokkoidi, rihmamainen, monisäikeinen, lamellaarinen ja sifonaalinen (kuva 44). – 46). Tallus yksisoluinen,

Riisi. 44. Kelta-vihreiden levien ulkonäkö: 1, 2 - Charatiopsis, 3 – Centritractus, 4 – ofiosytium
siirtomaa, monisoluinen ja ei-soluinen. Solukalvo on tiivistä, pektiinistä ja selluloosasta, joka koostuu tiiviisti limittäisistä osista tai kahdesta venttiilistä. Piidioksidia tai kalkkia kertyy kuoreen. Enimmäkseen kiinteät muodot. Yksisoluisten lajien joukossa on liikkuvia muotoja, joissa ei ole tiheää kuorta ja jotka on varustettu flagellalla, lobopodialla ja rhizopodialla.

Riisi. 45. Ksantokokin keltavihreän levien ulkonäkö: 1-3 - Botrydiopsis, 4 – tetrahedriella, 5 – Pseudostaurastrum, 6 – Goniochloris, 7, 8 – Bumilleriopsis
Useimmat kelta-vihreät – liikkumattomat organismit. Liikkuvilla yksilöillä liikkeet voidaan suorittaa käyttämällä flagellaa tai rhizopodiaa. Erimuotoisia soluja: pallomaisia, karan muotoisia, ellipsoidisia, sylinterimäisiä, tetraedrisä, sirpin muotoisia, päärynän muotoisia, munamaisia. Talluksen koot alkaen 0,5 – 1,5 µm ( Chloridella) halkaisijaltaan useita millimetrejä ( Botrydiopsis) (Kuva 45, 1 – 3) ja jopa kymmeniä senttejä pitkä ( Vosheria) (Kuva 46, 3).

Riisi. 46. Kelta-vihreiden levien ulkonäkö: 1 - Tribonema, 2 – heteropedia, 3 – Vosheria, osa filamenttia, jossa on oogoniumia ja anteridiumia
Suurin osa keltavihreistä lajeista on fototrofeja, mutta myös holotsoista ravintoa esiintyy bakteerien ja pienten levien nauttimisen kautta. Keltalevät ovat laajalle levinneitä makeissa vesissä. Ne ovat yleisiä myös maaperässä, vähemmän yleisiä meri- ja murtovesissä. Luokka sisältää aerobiontti-, plankton-, pohja- ja perifytoniset muodot. Epifyytit, epitsoiitit sekä solunsisäiset symbiontit alkueläinsoluissa.
Ulkoisesta rakenteesta riippumatta kelta-vihreäleväsolun sisäinen rakenne on sama. Protoplastissa on yleensä useita kellanvihreitä kloroplasteja, jotka ovat levymäisiä, kaukalomaisia, lamellisia, harvemmin nauhamaisia, tähtimäisiä tai kuppimaisia kokonaisia tai lohkoreunaisia. Väritys johtuu fukoksantiinin puuttumisesta, joka on vastuussa muiden okrofyyttien kultaisesta ja ruskeasta väristä. Niistä muista pigmenteistä β -karoteeni, vosheriaksantiini, diatoksantiini, diadinoksantiini, heteroksantiini. Liikkuvissa muodoissa punasilmäisyys tai stigma sijaitsee yleensä kloroplastin etupäässä. Harvalla lajilla on puoliksi upotettuja pyrenoideja. Solun ydin on yleensä yksi, pienikokoinen, mutta on lajeja, joissa on monitumaisia soluja. Joillakin lajeilla on yksi tai kaksi supistuvaa (sykkivää) vakuolia solun etuosassa.
Monadien edustajilla ja liikkuvilla vaiheilla (eläinporit ja sukusolut) on kaksi epätasa-arvoista siimat. Poikkeuksena ovat synzoosporit. Vosheri, jossa pinnalla on lukuisia sileitä, pituudeltaan hieman erilaisia siimoja. Lyhyt flagellum päättyy akroneemiin. Siimat kiinnittyvät soluun subaapisesti. Siittiössä Vosheri sivukiinnitys.
Lajeissa, joissa on ameboidi-, monadaalinen- ja palmelloidiorganisaatio, soluseinä puuttuu, niitä peittää vain sytoplasminen kalvo ja ne voivat helposti muuttaa muotoaan. Joskus "paljaita" soluja löytyy taloista, joiden seinät voidaan maalata ruskeiksi mangaani- ja rautasuoloilla. Suurimmalla osalla muotoja on soluseinä, joka koostuu kahdesta osasta. Selluloosa vallitsee soluseinän koostumuksessa, ja se sisältää myös polysakkarideja, jotka koostuvat pääasiassa glukoosista ja uronihapoista. Nuorissa soluissa kalvo on ohut, iän myötä se paksunee. Siihen voi kertyä rautasuoloja, joiden yhdisteet värjäävät sen ruskean ja punaisen eri sävyissä. Piidioksidia on useammin soluseinässä, mikä antaa sille kovuutta ja kiiltoa. Se voidaan myös peittää kalkilla ja muotoilla eri tavoin (piikit, solut, syyliä, syyliä, hampaat jne.) Kuoren uloskasvu voi muodostua kiinnittyneissä muodoissa – jalka kiinnityspohjalla.
Kelta-vihreälevien rihmamaisissa muodoissa, joissa on simpukoita, solukalvot hajoavat filamenttien hajottua H-muotoisiksi fragmenteiksi. Nämä fragmentit ovat tiiviisti yhdistettyjä kahden vierekkäisen solun kuoren puolikkaita (kuva 47). Filamenttien kasvun aikana kahden vierekkäisen tytärsolun soluseinän H-muotoinen fragmentti on upotettu emosolun kalvon kahden puolikkaan väliin. Tämän seurauksena jokainen tytärsolu on puoliksi peitetty emosolun vanhalla kalvolla ja puoliksi – vasta muodostettu kuori.

Riisi. 47. Kaavio poikittaisen väliseinän muodostumisesta kahden tytärsolun välille rihmamaisissa keltavihreissä levissä (A.A. Masyukin, 1993 mukaan): A- langan fragmentti; B- vyörenkaan asettaminen ja poikittaisen väliseinän muodostaminen kahden solun välille; SISÄÄN– simpukoiden solukalvojen kerrostaminen; G– kuoren hajoaminen H:n muotoisiin osiin
Liikkuvissa edustajissa on supistuvat tyhjiöt. Yleensä niitä on 1-2 per solu. Omalaatuisen rakenteen Golgi-laite. Diktyosomit ovat pieniä, sisältävät 3-7 säiliötä. Ydin on yksi, harvemmin niitä on useita; koenoottisissa lajeissa solut ovat aina moniytimiä.
jäljentäminen. Useimmille keltavihreille levälajeille on ominaista kasvullinen ja suvuton lisääntyminen.
Vegetatiivinen lisääntyminen toteutettu eri tavoilla: solun jakautuminen puoliksi, pesäkkeiden hajoaminen ja monisoluinen talli osiin. klo Vosheri muodostuu erityisiä pesäsilmuja.
klo suvuton lisääntyminen Erilaisia itiöitä voi muodostua: amoeboideja, zoosporeja, synzoosporeja, autoitiöitä, hemisoosporeja, hemiautosporeja, aplanosporeja. Zoosporit ovat "alastomia" ja yleensä päärynän muotoisia.
seksuaalinen prosessi- isogamia, heterogamia ja oogamia - kuvattu muutamassa edustajassa. klo Tribonemes Sukusolut ovat kooltaan samanlaisia, mutta käyttäytyvät erilaiset - tämä on isogamiaa. klo Vosheri Oogamiaa havaitaan: lankoihin muodostuu naarassukusolujen astioita – oogonia ja uros – anteridia.
SISÄÄN epäsuotuisat olosuhteet havaitaan kystin muodostumista. Kystat (statosporit) ovat endogeenisiä, yksitumaisia, harvoin moniytimiä. Niiden seinämä sisältää usein piidioksidia ja koostuu kahdesta epätasaisesta tai harvemmin yhtä suuresta osasta.
Systematiikka.
XIX vuoden lopussa – 1900-luvun alku Viherleväosastolle sijoitettiin useita keltavihreäleväsuvuja, jotka liittyivät ensisijaisesti tallin väriin ja morfologiseen samankaltaisuuteen. Keltaisia vihreitä pidetään tällä hetkellä okrofyyttiluokkaan kuuluvana luokkana.
Noin 450 tiedossa nykyaikaiset lajit luokka Keltavihreät levät, jotka on ryhmitelty neljään luokkaan: Botridia, Mishococcus, Tribonem ja Wocheriaceae. Tilausten luokittelu perustuu talluksen erilaistumistyyppiin ja elinkaaren ominaisuuksiin.
Tilaa Botridiae - Botrydiales. Luokka sisältää lajeja, joilla on sifonaalinen talluksen erilaistuminen, jossa ei ole oogaamia sukupuoliprosessia.
Suku Botridium elää maaperässä ja näyttää muutaman millimetrin kokoisilta vihreiltä kuplilta, joihin on kiinnitetty värittömiä risoideja. Tallus on sifonaalinen, sisältää lukuisia ytimiä ja plastideja. Kuori on monikerroksinen, sen päälle voidaan kerrostaa kalkkia. Lisääntyminen on suvutonta biflagellate zoosporien avulla, kun taas rakon koko sisältö hajoaa yksitumaisiksi fragmenteiksi. Kosteuden puutteessa se lisääntyy aplanosporien avulla tai muodostaa paksuseinäisiä kystoja. Joissakin tapauksissa koko virtsarakon sisältö menee yhden suuren kystan muodostumiseen. Muissa tapauksissa kystat muodostuvat risoideihin, joissa virtsarakon sisältö ensin kulkeutuu. Kystat itävät joko suoraan uuteen tallukseen tai muodostavat zoosporeja. Seksuaalinen prosessi on iso- ja heterogamia. Tsygootti itää välittömästi, ilman lepojaksoa. Yleisiä ja laajalle levinneitä lajeja maa-elinympäristöissä, joita tavataan purojen, lampien rannoilla tai maaperällä, jossa ei ole kasvillisuutta.
Tilaa Mischococcal–Mishococcaleja. Yksisoluiset tai koloniaaliset edustajat, joilla on kokkoidityyppinen talluksen erilaistuminen.
Suku Charatiopsis sisältää yksisoluiset kiinnittyneet muodot. Lisääntyessään se muodostaa zoosporeja, aplanosporeja ja paksuseinäisiä kystoja (Kuva 44, 1-2).
Suku ofiosytium(Kuva 44, 4) on pitkänomaiset lieriömäiset kennot, jotka voivat olla suoria, taivutettuja tai spiraalimaisesti kierrettyjä ja joissa voi olla piikki. Soluseinä koostuu kahdesta epätasaisesta osasta, joista suurin osa osallistuu solujen kasvuun, pienempi osa on pysyvä ja korkin muotoinen. Yksisoluinen ja siirtomaalaji, vapaasti elävä tai kiinnittynyt alustaan pienellä varrella. Ne lisääntyvät zoosporien ja aplanosporien avulla, kystat löytyvät. He elävät makeissa vesissä.
Suku Mischococcus muodostaa puumaisia kiinnittyneitä pesäkkeitä. Haaroittuminen on kaksijakoista ja tetrakotomista. Solut sijaitsevat 2 tai 4 pesäkkeen limahaarojen yläosissa. Solut ovat pallomaisia tai soikeita, ja niissä on ohut tai paksu soluseinä. Joskus soluseinä on kiiltävä ja ruskea, koska se on kyllästetty rautasuoloilla. Nuori yksisoluisia organismeja jossa on limainen, kiekon muotoinen pohja, joka toimii kiinnityksen tukipisteenä. Itiöiden vapautumisen jälkeen emosolun protoplasti muuttuu hyytelöksi ja venyy, pituus on 6 kertaa suurempi kuin leveys, ja siten syntyy lieriömäinen jalka. Emosolun tyhjä soluseinä tulee aina kantasolun pohjaksi. Aseksuaalinen lisääntyminen zoosporien ja autosporien avulla. Autoitiöt ovat kiinnittyneet limakalvon varren yläreunaan. Seuraavat solujakautumat toistavat prosessin ja tuottavat puun kaltaisen pesäkkeen. seksuaalinen prosessi – isogamia. Ne elävät pienissä makeissa vesistöissä rihmamaisten levien epifyytteinä. Tunnettu Keski-Euroopassa ja Aasiassa.
Tilaa Tribonemaceae - Tribonematales. Edustajilla on rihmamainen, monisäikeinen, väärän kudoksen ja kudostyyppinen talluksen erilaistuminen. Soluseinät ovat joko H-muotoisia päällekkäisiä osia tai kiinteät.
Suku Tribonema- haarautumattomat kierteet (kuva 46, 1). Solut ovat sylinterin tai tynnyrin muotoisia. Soluseinä koostuu kahdesta puolikkaasta, joiden reunat ovat päällekkäin solun keskellä. Kuoret ovat usein kerroksittain. Lankojen palaset päättyvät aina tyhjiin H-muotoisten kuorikappaleiden puolikkaisiin, jotka näyttävät haarukalta. Soluissa on useita kellertävänvihreitä plastideja, pyrenoideja ei ole. Lisääntyminen on vegetatiivista (filamenttien sirpaloituminen), aseksuaalista (zoosporien ja aplanosporien kautta) ja sukupuolista (isogamia), ja aplanosporeja tuotetaan useammin kuin zoosporeja. Voi muodostaa akinetteja. Ne elävät makeissa vesissä, joissa ne kehittyvät erityisen runsaasti kylmänä vuodenaikana.
Tilaa Vosherievye - Vaucheriales. Kaikilla edustajilla on sifonaalinen tallus, oogaminen seksuaalinen prosessi ja synzoosporit.
Suku Vosheria(kuvio 46, 3) on tallus, jolla on ei-solurakenne; sen tallus saavuttaa useiden senttimetrien pituuden, kiinnittyy alustaan värittömän risoidin avulla. Langoissa ei ole väliseiniä, suurin osa talluksesta on tyhjiössä, ja lukuisat ytimet ja plastidet sijaitsevat sytoplasman reunalla. Langat, joissa on apikaalinen kasvu ja harvinainen sivuhaaroitus. Väliseinät muodostuvat, kun talli on vaurioitunut ja erottamaan sukuelimet. Aseksuaalista lisääntymistä suorittavat aplanosporit, synzoosporit, akinetit. Synzoosporit muodostuvat yksi kerrallaan zoosporangiumissa, joka on erotettu vegetatiivisista soluista filamentin päässä olevalla väliseinällä. Eläinporit ovat monitumaisia ja monisiimaisia. Seksuaalinen prosessi on oogamiaa. Tsygootti on peitetty paksulla kuorella ja kasvaa lepotilan jälkeen uudeksi tallukseksi.
Erilaisia Vosheri levinnyt laajalti makeissa, murto- ja merivesissä sekä maalla olevissa elinympäristöissä. Niitä löytyy kaikilla mantereilla, myös Etelämantereella. Ne muodostavat ruohonvihreitä tai tummanvihreitä toisiinsa kietoutuneita massoja - niin sanottuja mattoja, sileitä, hiipiviä tai tyynynmuotoisia. Vesi-, puolivesi-, maanpäälliset muodot. Ne elävät erilaisissa elinympäristöissä: merissä, suistoissa, suistoissa, suolamaissa, mangrovemetsissä, puroissa, kanavissa, järvissä, lammissa, peltomailla ja soilla.
Merkitysheterokont levät
Heterocont-leväosastolta ruskealevällä on suurin merkitys luonnollisille ekosysteemeille ja ihmisille.
Ruskeat levät - tärkein orgaanisen aineen lähde merten rannikkovyöhykkeellä. Niiden biomassa lauhkean ja subpolaarisen vyöhykkeen merissä voi olla useita kymmeniä kiloja neliömetriä kohti. Ruskealeväpaksut tarjoavat olosuhteet monien rannikkoeläinten ja muiden levien ruokimiselle ja lisääntymiselle. Charles Darwin tarkkailee ruskealevää Etelä-Amerikan rannikolla Makrocystis, kirjoitti: "Voin vain verrata näitä eteläisen pallonpuoliskon valtavia vedenalaisia metsiä trooppisten alueiden maanpäällisiin metsiin. Ja silti, jos jossain maassa tuhoutuisi metsä, en usko, että edes suunnilleen sama määrä eläinlajeja kuolisi kuin tämän levän tuhoutuessa.
Ruskealevän paksut palvelevat ruokapaikka,suojaa ja kasvatus monia eläimiä. Kuvaannollisesti sanottuna ruskeat levät tarjoavat muille vesieliöille "pöydän, suojan ja lastenhuoneen".
Ruskealevää käytetään laajalti ihmisillä. He ovat rikkaita jodi ja muita mikroravinteita. Kaakkois-Aasian kansat käyttävät niitä perinteisesti ruoaksi, erityisesti rakkolevälajin edustajia, joista valmistetaan monenlaisia ruokia. Syö ateria ruskealevistä valmistettu, lisää karjan tuottavuutta; Samalla munien ja maidon jodipitoisuus kasvaa.
Ruskeista levistä vastaanottaaalginaatit- algiinihapon suolat. Alginaatteja käytetään laajasti eri teollisuudenaloilla. Nämä ovat myrkyttömiä yhdisteitä, joilla on kolloidisia ominaisuuksia, joten niitä käytetään laajalti elintarvike- ja lääketeollisuudessa. Algiinihappo ja sen suolat kykenevät 200-300-kertaisesti imemään vettä muodostaen geelejä, joille on ominaista korkea haponkestävyys. Elintarviketeollisuudessa niitä käytetään pääasiassa emulgointiaineina, stabilointiaineina, hyytelöimis- ja vettä pidättävinä komponentteina. Esimerkiksi kuivajauhenatriumalginaattia käytetään jauhettujen ja briketoitujen liukenevien tuotteiden (kahvi, tee, maitojauhe, hyytelö jne.) valmistuksessa niiden nopeaan liukenemiseen. Vesipitoiset liuokset alginaatteja käytetään liha- ja kalatuotteiden pakastamiseen. Jopa 30 % maailmassa tuotettujen alginaattien kokonaismäärästä menee elintarviketeollisuuteen.
Tekstiili- ja massa- ja paperiteollisuudessa alginaatteja käytetään maalin sakeuttamiseen ja niiden sidoksen lujuuden lisäämiseen pohjan kanssa. Kankaiden kyllästäminen joillakin alginaateilla antaa niille suojaavia ominaisuuksia: vedenpitävyyttä, haponkestävyyttä ja lisää mekaanista lujuutta. Keinotekoisen silkin valmistamiseen käytetään lukuisia algiinihappojen suoloja. Toisen maailmansodan aikana Yhdysvalloissa ja Englannissa valmistettiin algiinihaposta ja sen suoloista suuri määrä naamiointikankaita ja verkkoja asuin- ja teollisuusrakennuksiin.
Alginaatteja käytetään metallurgiassa: valimossa ne parantavat valumaan laatua. Algiinihapon suoloja käytetään sähköhitsauksen elektrodien valmistuksessa, mikä mahdollistaa laadukkaampien hitsausten valmistamisen. Alginaatteja käytetään myös muovien, synteettisten kuitujen, pinnoitteiden ja säänkestävien rakennusmateriaalien valmistuksessa. Niitä käytetään korkealaatuisten koneiden voiteluaineiden valmistukseen. Radioelektroniikassa alginaatit toimivat sideaineena korkealaatuisten ferriittien valmistuksessa.
Suurin sovellus löytyy vesiliukoisesta natriumalginaatista, joka pystyy muodostamaan viskoosisia liuoksia. Sitä käytetään laajasti erilaisten liuosten ja suspensioiden stabilointiin. Pienen määrän natriumalginaattia lisäämällä elintarvikkeisiin - säilykkeisiin, jäätelöön - niiden laatu paranee. Siitä valmistetaan myös koristekosmetiikkaa, voiteita ja naamioita hajuvesiteollisuudessa.
Lääketeollisuudessa alginaatteja käytetään tablettien, pillereiden päällystämiseen, erilaisten voiteiden ja tahnojen komponenttipohjana, lääkekantajageeleinä sekä liukenevien kirurgisten ompeleiden valmistuksessa. Lääketieteessä kalsiumalginaattia käytetään hemostaattisena aineena ja radionuklideja (esimerkiksi strontiumia) poistavana sorbenttina. Alginaattien vuosituotanto maailmassa ylittää 20 tuhatta tonnia.
Toinen tärkeä ruskealevistä peräisin oleva aine on kuusiarvoinen alkoholimannitoli. Mannitolia käytetään sokerinkorvikkeena diabeetikoille. Lisäksi sitä voidaan käyttää plasman korvikkeena veren säilönnässä. Sitä käytetään tablettien valmistukseen lääketeollisuudessa. Mannitolia käytetään myös mm synteettisten hartsien, maalien, paperin, räjähteiden, nahkasidosten tuotanto.
fukoidaanit, ruskealevistä saadut ovat tehokkaita antikoagulantteja, jopa hepariinia aktiivisempia. Niiden käyttöä kasvainlääkkeiden ja viruslääkkeiden valmistukseen pidetään lupaavana. Loppujen lopuksi, jopa pienimmällä pitoisuudella, fukoidaanit voivat estää virusten kiinnittymisen solujen pintaan. Fukoidaaneilla on myös kyky muodostaa poikkeuksellisen vahvaa ja viskoosia limaa, jota käytetään stabiilien emulsioiden ja suspensioiden valmistukseen.
Monia maailman maita viime vuosina valtannut energiakriisi on johtanut tarpeeseen etsiä uusia ei-perinteisiä energialähteitä. Joten Yhdysvalloissa tätä tarkoitusta varten tutkitaan mahdollisuutta kasvattaa jättiläisleväleviä. Makrocystis jonka jälkeen se prosessoidaan metaaniksi. On laskettu, että tämän levän 400 km 2:n alueelta voidaan saada 620 miljoonaa m 3 metaania.
Heterokonttilevät luokista Golden, Yellow-green, Sinura, Rafidophyte ja Eustigma, joita edustavat pääasiassa mikroskooppiset organismit, ovat yleisiä makeissa vesistöissä kaikilla maapallon ilmastovyöhykkeillä, mutta ovat yleisempiä lauhkeilla leveysasteilla. Kultalevien joukossa on lajeja, jotka elävät merissä ja suolajärvissä, ja hyvin harvat saastuneissa vesissä. Kultalevät saavuttavat huippukehityksensä kylmänä vuodenaikana: ne hallitsevat planktonissa aikaisin keväällä, myöhään syksyllä ja talvella. Tällä hetkellä niillä on merkittävä rooli alkutuotannon tuottajina ja ne toimivat eläinplankton-organismien ravinnoksi.
Jotkut kultalevät esimerkiksi Uroglena Ja Dinobrion, kehittyvät massamäärinä, ne voivat aiheuttaa vesikukintoja. Ne vapauttavat aldehydejä ja ketoneja, jotka voivat antaa vedelle epämiellyttävän hajun ja maun, a Uroglena– kalalle myrkyllisiä rasvahappoja.
Rafid-levät ovat laajalti edustettuina pääosin happaman pH:n makean vesistön planktonissa, erityisesti sfagnum-soissa, harvemmin suurissa järvissä. Makeissa vesistöissä voi muodostua paikallisia "kukintoja". Goniostomum. Rafid-leviä esiintyy myös suolattomissa merenlahdeissa ja lätäköissä merenrannalla sekä avomerellä. Rannikon merivesien massojen kehittyessä ne aiheuttavat myrkyllistä veden "kukintoa". Joten Kanadan rannikolla "kukinnan" aikana Heterosigma-rafid-levän solujen pitoisuus voi nousta 30 miljoonaan litrassa. Rapidlevien puhkeaminen johtaa usein "punaisten vuorovesien" kehittymiseen, jotka liittyvät kalojen kuolemaan. Tällaisten "punaveden" syy voi olla synnytyslajit Hattonella, Olistodiscus, Heterosigma ja Fibrocapsa.
Makeissa vesistöissä massoittain kehittyneet sinuralevät voivat aiheuttaa veteen epämiellyttävän hajun ( Sinura). Feotamnialeviä esiintyy pysähtyneissä ja hitaasti virtaavissa makeissa vesistöissä, joissa ne asettuvat epifyyttisesti rihmaleville.
Eustigma-levää esiintyy vain makeassa vedessä tai maaperässä.
Keltaleviä on levinnyt kaikille mantereille, ne elävät pääasiassa makeassa vedessä ja maaperässä sekä maa-, murtovesi- ja meriympäristöissä. Keltalevät elävät puhtaissa ja saastuneissa vesissä, joiden pH-arvot vaihtelevat: ne voivat elää sekä happamissa että emäksissä. Niitä esiintyy pääasiassa puhtaissa makean veden säiliöissä, harvemmin merissä ja murtovesissä, mieluummin maltillisia lämpötiloja, kehittyvät useammin keväällä ja syksyllä, vaikka on lajeja, joita esiintyy kaikkina vuodenaikoina, myös talvella. Useimmiten ne löytyvät lankaryhmistä ja korkeammista pensaikkoista vesikasveja jokien, lampien, järvien ja tekoaltaiden rannikkoalueella.
Suurin osa keltavihreistä on vapaasti eläviä muotoja, mutta alkueläinsoluissa on myös solunsisäisiä symbiontteja - zooksanteeleja. Meren lajien kloroplastit muodostavat mielenkiintoisen solunsisäisen symbioosin Vosheri simpukan kanssa Elisio. Yhdeksän kuukauden ajan tämä nilviäinen pystyy kiinnittämään fotoautotrofisesti hiilidioksidia viljelmässä. Tämä on pisin tämän tyyppinen symbioosi, kun symbioottinen plastidi on suorassa kosketuksessa eläimen sytoplasmaan. Luonnossa nilviäisten toukat ruokkivat lankoja. Vosheri. Fagosytoosin seurauksena leväkloroplastit pääsevät nilviäisten epiteelisolujen sytoplasmaan. Tämän prosessin aikana kloroplastikalvo muuttuu kolmikerroksiseksi, ja yksi CES:n (kloroplastin endoplasminen retikulum) ulkokalvo menetetään. Tämä ilmiö on hyvä todiste siitä, että evoluution aikana kolme kalvoa sisältäviä kloroplasteja voi syntyä sekundaarisen symbiogeneesin seurauksena kalvojen häviämisen seurauksena.
Keltavihreät, kultaiset ja muut heterokonttilevät ovat hapen ja orgaanisten aineiden tuottajia; ne ovat osa ravintoketjua. Heterokontofyytit osallistuvat saastuneiden vesien ja maaperän itsepuhdistumiseen, sapropeelin muodostumiseen, orgaanisen aineksen kertymiseen maaperään, mikä vaikuttaa sen hedelmällisyyteen. Niitä käytetään edustavina organismeina määritettäessä veden saastumisen tilaa; keltavihreät levät ovat osa jäteveden käsittelyyn käytettyä mikro-organismikompleksia.
Kontrollikysymykset
Nimeä ruskealevän rakenteen ominaispiirteet.
Ruskealeväthallin rakenteelliset piirteet.
Kuinka ruskeat levät lisääntyvät? Mitä ovat monosporit, tetrasporit ja zoospores, isogamia, heterogamia ja oogamia?
Mitkä ovat ruskealevien elinkaaret? Fucus- ja rakkolevän lisääntyminen.
Nimeä ruskealevälajien tunnusmerkit ja tyypilliset edustajat.
Missä elinympäristöissä ruskealeviä löytyy? Mikä on niiden merkitys luonnossa.
Ruskealevien taloudellinen merkitys.
Nimeä kultalevän rakenteen ominaispiirteet ja tyypilliset edustajat.
Mitä pigmenttejä ja ravintotyyppejä kultalevistä tunnetaan?
Kultalevien lisääntyminen ja ekologia.
Nimeä kelta-vihreiden levien tunnusmerkit ja tyypilliset edustajat.
Mitä pigmenttejä ja ravintotyyppejä keltalevissä tunnetaan?
Miten keltavihreät lisääntyvät? Sukupuolisen lisääntymisen tyypit: isogamia, heterogamia ja oogamia?
Nimeä sinilevän rakenteen ominaispiirteet ja tyypilliset edustajat.
Sinuurilevien lisääntyminen ja ekologia.
Nimeä feotamnialevien rakenteen ominaispiirteet ja tyypilliset edustajat.
Feotamnialevien lisääntyminen ja ekologia.
Nimeä rafidofyyttilevien rakenteen yleiset piirteet ja tyypilliset edustajat.
Rafidophyte levien lisääntyminen ja ekologia.
Mitkä ovat Eustigma-levien rakenteen ja tyypillisten edustajien yhteiset piirteet.
Eustigma-levien lisääntyminen ja ekologia.
Heterokontlevien arvo luonnollisissa ekosysteemeissä.
MERILEVÄ
Yleiset luonteenpiirteet
Levät ovat suuri joukko alempia kasvejany - mikroskooppisesti pienistä yksisoluisista organismeistamonisoluisia jättiläisiä. Termi "levät" ei ole järjestelmämatemaattinen yksikkö. Nimen alla "levät" yhdistyvätuseat eri alkuperää olevien alempien kasvien systemaattiset jaot, jotka on luonnehdittu samalla tavallaelämä ja autotrofinen ravitsemustyyppi. Kuten nimestä voi päätellä,leville on yleensä ominaista vesielämys.
Levien runko, kuten muutkin alemmat kasvit, ontallus - talli, ei leikattu juureksi, varsija lehtiä, ja useimmissa lajeissa on yksinkertainen anatominen rakenne. Levät sisältävät klorofylliä soluissaanfotosyntetisoida. Rakennesolut, elimet, lisääntymismenetelmät ovat erilaisia yksittäisillä edustajillatämän tehdasosastojen ryhmän edustajat.
Useimpien levien soluissa on selluloosakalvo; on soluja, joiden sisältöä ympäröi vain ohut rajakalvo. Levien soluseinät ovat rakenteeltaan ja monimuotoisia kemiallinen koostumus. Shell-pohjainenon proteiini- hiilihydraattikompleksi. Kuorelle on ominaista heterogeenisyys, kerrostuminen. Kerrokset eroavat toisistaanpaksuus, tiheys ja kemiallinen koostumus. Usein kuoret kyllästetään orgaaniset yhdisteet(ligniini ja kutiini).
Kuoressa on erityisiä reikiä - huokosia. Huokosten lisäksi monien levien solukalvot on varustettu erilaisilla kasvaimilla.stami - harjakset, piikit ja suomut.
Solun protoplastissa erotetaan ydin ja sytoplasma. Tee kipuaUseimmilla solun levillä on vain yksi ydin, mutta joskusNiitä on 2, 3 tai enemmän. Sinilevän soluista puuttuu hyvin muodostunut ydin. Ytimen muoto, koko ja sijainti solussavaihtelevat suuresti eri levien välillä. Levän ytimessämuodostuvat samat rakenteet kuin muiden kasvien ytimiin: kuori,ydinmahla, nukleoli, kromatiinisulkeumat.
Sytoplasma koostuu pääaineesta (strooma) ja siihen upotetuista kappaleista (organelleista). Liiman erottuva ominaisuuslevävirta on endoplasmisen retikulumin heikko kehitys.
Tietyissä organelleissa - kloroplasteissa (kutsutaan kromomatoforit) ovat runsaita proteiiniaineita,kutsutaan pyrenoideiksi. Pyrenoidia ympäröi vuori, joka on renkaan tai yksittäisten levyjen muodossa, yleensä tärkkelyspitoinenluonto. Kloroplastit sisältävät vihreää pigmenttiä klorofylliäolemassa useissa muodoissa.
Klorofyllin lisäksi levät sisältävät muitakin pigmenttejä, jotkajotka usein peittävät läsnäolollaan veden vihreän värin.kasva aikuiseksi. Ominaisimmat ja useimmin kohtaamat sikamiehetlevät, paitsi klorofylli (vihreä), karoteeni (keltainen-oranssi) ja ksantofylli (keltainen), ovat fykosyaniinia, fyko-erytriini ja fukoksantiini. Fykosyaanipigmentti, liukenee veteen,värit levä sininen (sinilevä);fykoerytriini, joka myös liukenee veteen, antaa punaisen värin(punalevä); fukoksantiini aiheuttaa ruskean värin(ruskolevät).
Levät, joilla on pigmenttejä, antavat substraatin, jollene asettuvat, eri väritys. Esimerkiksi Etelämantereella Neuvostoliiton tutkijat löysivät kolme eriväristä järveävesi: sininen, vihreä ja punainen. Näiden järvien veden väri johtuihyvin pienten, vastaavan väristen levien esiintyminen niissä.Grönlannin jäärannat monin paikoin kevään alkaessasen lämpeneminen muuttuu punaiseksi (veriseksi) väriksi, mikä selittyy punaisten levien lisääntymiselläkaino.
Punaisenmeren nimi johtuu sen kasvusta suurenatässä meressä olevien levien määrä, joilla on punertava sävy.Levän eri väreillä on mukautuva arvoei. Päivänvalo syvälle veteen upotettuihin leviintulee aina muunnetussa koostumuksessa. Vesi on suhteellisen hyvää.siirtää sinisiä ja vihreitä ja imee voimakkaasti punaisia ja keltaisiasäteet. Suurissa syvyyksissä vihreiden säteiden pysyminen yhdelläklorofylli on huono, punainen tulee apuunpigmentti, joka imee helposti vihreitä säteitä. Siksi punainentai karmiininpunainen, levät ovat yleisiä suurissa syvyyksissämeret.
Levät ovat muodoltaan ja kooltaan hyvin erilaisia. Tavata-On olemassa mikroskooppisesti pieniä ja valtavia kokoja (jopa 50 m ja enemmän) yksisoluisia ja monisoluisia muotoja. Siirtymävaiheen niiden välillä vallitsevat siirtomaalevät.Pesäkkeet koostuvat useista löyhästi toisiinsa liittyvistä homogeenisistasoluja. Levien koon kasvaessa jotkutheidän ruumiinsa paratiisi erilaistuminen. Esimerkiksi ruskeissa levissäkasvaa suureksi, tallus voimakkaasti leikattu. Tällaiset levät kiinnitetään alustaan ohuilla värittömillä langoilla,joita kutsutaan risoideiksi.
On yksisoluisia leviä, joihin keho saavuttaasuuret koot 0,5 m ja enemmän. Niitä on suuri määräytimet ja kromatoforit. Ulkoisesti tällaisten levien ruumis on hajotettumutta sisällä ei ole osioita, eli se on yksisoluinen. Esimerkki tällaisesta jättimäisestä yksisoluisesta levistä onelävä merellinen caulerpa.
Levien lisääntyminen voi olla vegetatiivista, aseksuaalista ja seksuaalista. Vegetatiivinen lisääntyminen tapahtuu osissaleväkappaleita. Aseksuaalinen lisääntyminen tapahtuu zoosporien, harvemmin itiöiden, muodostumisen vuoksi. Seksuaalinen prosessi eri tyypeissälevää edustavat: isogamia, heterogamia, oogamia jaautogamia.
Joissakin paremmin organisoituneissa levissä (punainen,ruskea) seksuaaliset ja aseksuaaliset sukupolvet vaihtuvat.
Leveille on ominaista suuri monimuotoisuus. Kenraaliniiden lajien lukumäärä on yli 20 tuhatta. Levien luokitus on erittäinmonimutkainen eikä ole vielä valmis. Lajit yhdistetään suuremmiksi systemaattisiksi yksiköiksi - suvuiksi, perheiksistva, tilaukset, luokat, osastot (tyypit).
Koko levävalikoima yhdistetään yleensä 6 ... 10 suureen osastoon (tyyppiin), joiden edustajat eroavat rakenteeltaanny, lisääntymismenetelmä, alkuperä, mutta useimmiten ennenNäiden osastojen edustajat eroavat toisistaan väriltään.
Tässä oppikirjassa tarkastellaan 6 leväosastoa: sini-vihreä, multiflagellate, piilevät, vihreä, ruskea ja punainennye.
Levät ovat hyvin ikivanha alkuperä, jossapaljon on vielä epäselvää. Ei ole yhtä näkemystä leväjakojen välisestä suhteesta. Jotkut heidän edustajistaanilmeisesti peräisin yksinkertaisemmista organismeista, kuten flagellateista, ja levät itse ovat joidenkin muiden aikojen esi-isiäkierretyt kasvit (sienet, sammalet).
Modernin monipuolisen levämaailman edustajatniiden alkuperä on erilainen, ne ilmestyivät eri geologisina aikakausina. Vanhimmat ovatsinilevät.
Levien evoluutio eteni liikkuvista muodoista liikkumattomiksi.Alkukantaisempia ja muinaisempia leväryhmiä pidetäänne, jotka viettävät elämänsä liikkuvassa tilassa; enemmän organisointiaKutsutuille leville on ominaista kiinteä elämäntapa. Polkulevien evoluutio tuli alkueläimistä - yksisoluisista mikroistaskooppisen pienistä muodoista monisoluisiin kompleksimuotoihin.
Kuten jo mainittiin, suurin osa levistäelää vesiympäristössä. Jotkut niistä kasvavat valtamerten suolaisessa vedessä.ja meret, muut - joissa, lammissa ja makean veden järvissä. Kipuvaikutus levien leviämiseen vesistöjen syvyyksissäantaa valoa, joten veden pintakerroksissa määräleviä on aina enemmän. Merilevä punainen ja ruskeamuodostavat usein valtavia vedenalaisia paksuja, jotka kattavat kymmeniä kilometrejä. Elinympäristöstä riippuen levätjaetaan kahteen suureen ryhmään: pohjaeliöön ja planktoniin.
Pohjalevät eli pohjalevät elävät kiinnittyessäänpohja- tai vedenalaiset esineet ja muodostavat pääasiassa paksujarannikkoalueella. Planktonlevät elävät vedessäripustettuna, ei kiinnitetty pohjaan, ne voidaan siirtääliikkua veden liikkeen mukana.
Levät elävät paitsi vedessä, myös pinnallamaaperässä, maaperässä, puissa, paaluissa, mutta aina kosteanapaikoissa. Usein, varsinkin aikaisin keväällä, maaperä "kukkii",tai "muuttu vihreäksi", mikä selittyy jättimäisen colin kehittymisellämikroskooppisesti pieniä leviä. maaperän "kukkiminen".levää esiintyy eri alueilla Neuvostoliitto - pohjoisessa, aroissa ja jopa autiomaassa. Kasvaessa suurissa määrissä, ne luovat olosuhteet bakteerien ja muiden kehittymisellemikro-organismeja.
Levillä on hyvin tärkeä luonnossa ja laajalti käytössäjoita henkilö kutsuu.
Autotrofisina kasveina ne käsittelevät valtavia koliamineraalien määrä ja hiilidioksidi orgaaniseksi aineeksi. Levät luovat valtavia ravintovarastoja eläimellemerien, valtamerten ja makean veden altaiden maailma. Joten 1 ha leväpehmikistä voi tuottaa satoa, joka vastaa 100 tonnia raakaa tai 10 tonnia kuivaa.massat.
Levät tuottavat valtavan määrän happea.
Kalastuksen kehitys liittyy erottamattomasti leviin. Imeyttämällä paljon hiilidioksidia ja vapauttamalla happea, leviäpuhdistaa säiliöitä.
Rannikkomaissa (Englanti, Ranska, Norja, Irlantijne.), erityisesti Japanissa, leviä syödään laajalti(merilevä jne.) ja karjan rehuksi (raaka-, kuiva- ja säilörehu).kylpyhuone). Joitakin leviä käytetään lannoitteinakentät.
Monet levät keräävät suuria määriä jodia ja bromia. Jodia on levien tuhkassa noin 0,2 % kuivamassasta.Levät ovat usein pääasiallinen jodin lähde. Ruoansulatus, punaiset ja ruskeat levät, saavat arvokasta ainetta - agar-agaria,jota käytetään mikrobiologiassa ravintoalustanamikro-organismien viljelyyn sekä makeisissamarmeladin valmistusteollisuus jne.
Muinaisilla geologisilla aikakausilla piilevät yhdessäpiidioksidin kanssa muodostui sedimenttikivi - tripoli, jokakäytetään dynamiitin, tiilien valmistukseen, kiillotukseenesineitä jne.
Makeissa vesistöissä levät osallistuvat sapropeen muodostumiseenla tai orgaaninen liete. Sapropeli sisältää suuren määränorgaanisessa aineessa ja sitä käytetään usein mutahoitoon.Sapropeli, joka sisältää suuren määrän kalsiumsuolojaioneja, fosforia, rautaa, voidaan käyttää maatalouden rehuissataloudelliset eläimet.
Levät, erityisesti merilevät, voivat myös olla haitallisia, kunsuuria määriä peittävät aluksen vedenalaiset osat. klolevien massakuoleminen aiheuttaa vesivahinkoja ja mmnäin ollen kalojen kuolema.
Sinilevien jako
Sinilevät eli syanidi ovatalkukantaisin ja alkuperältään vanhineliöt. Perustuu paleobotanisiin tietoihinOn surkeaa, että nykyaikaiset sinilevät eivät eroa paljon toisistaanfossiilisista esivanhemmistaan. Useimmissa tapauksissa niitä edustavat yksisoluiset muodot, vaikka on myös monisoluisiasiirtokunnassa kerätyt nye-lomakkeet.
Kuten tämän leväosaston nimi osoittaa, heilletyypillinen sinivihreä väri eri sävyillä riippuensillat pigmenttien suhteen - klorofylli, karoteeni, fyko-syaniini ja fykoerytriini.
Sinilevän solukalvo koostuu pektiinistävyh aineita ja on ulkopuolelta peitetty limakalvolla. Heidän häkissäänkah:lta puuttuu morfologisesti erottuva ydin ja kromatoforit,sytoplasma on kyllästetty pigmenteillä, ja siksi sitä kutsutaan kromatoksiplasma.
Assimilaatioprosessissa tavanomaisen kasvitärkkelyksen sijaanmuodostuu pientä hiilihydraattiglykogeenia (eläintärkkelystä).
Sinilevät lisääntyvät hyvin nopeasti, yksinkertaisellasolujen puolittaminen. Heidän seksuaalinen lisääntymisprosessinsapoissa.
Sinivihreän veden rakenteen ja solujakautumisen luonteen mukaankasvut ovat samanlaisia kuin bakteerit. Heillä, kuten bakteereilla, ei ole selkeästi määriteltyä ydintä, niillä ei ole siimakehitysvaiheita.olla olemassa. Joissakin sinilevissä (rihmamaiset)Vähentyminen tapahtuu alueilla, joihin nämä levät hajoavat. Tällaisia alueita kutsutaan hormogoniaksi. Kun epäsuotuisamiellyttävissä olosuhteissa tavallisista soluista muodostuu itiöitä, jotkapeitetty paksulla vaipalla. Se suojaa sisältöäepäsuotuisista olosuhteista, minkä ansiosta levät säilyvätelinkelpoisuutta pitkään. Alkuhetkelläsuotuisissa olosuhteissa itiöt itävät ja synnyttävät uuttasolu.
Sinilevät elävät pääasiassa makeassa vedessä.vedet - lammet, järvet, joet, mutta myös merissä,maan pinnalla, kivillä. Sinilevä voielää kuin matalat lämpötilat lumella ja jäällä sekä korkeallalämpötiloissa (jopa 80 °C) kuumissa lähteissä. Kuoleman jälkeenleväsolumassa likaisen vihertävän väristen hiutaleiden muodossakelluu veden pinnalle.
Liiman muinainen alkuperä, erilaistumaton rakennevirta, muodostuneen ytimen ja sukupuoliprosessin puuttuminen, lisääntyminenyksinkertaisen solunjakautumisen kautta kyky muodostaa itiöitä -kaikki nämä ominaisuudet viittaavat sinivihreän primitiivisyyteenlevät. yksinkertaistettu omalla tavallaantietylle rakenteelle ne ovat merkittävästieroaa muista levistäHän ja lähimpänä seistä bakteereja.
Sinilevällä on hyvä sopeutumiskykyvastustuskyky erilaisille olosuhteillekuoppia ympäristöön, mikä onedistänyt niiden säilymistätähän päivään asti ilman paljonmuutoksia.
Sinivihreän vedyn jakosli yhdistää noin 1400 widov. Tämän osaston edustajat voivat olla tällaista vettäkasvoi kuten chroococcus, oscillatoria, nostoc jne.
Chroococcus- yksisoluinen levä Sharonäkyvä muoto, joskus nämä levät muodostavat ikään kuin pesäkkeitä.Usein tällaiset levät kerätään 2 ... 4 ryhmiin erotettuinaohut väliseinä ja sitä ympäröi tavallinen melko paksu limapaksu kerros.
Chroococcus on laajalti levinnyt suoissa vesirotujen keskuudessa.varjoja, kuoppien ja mudan välissä.
Oskilloiva- filamenttinen sinivihreälevät, jotka ovat yleisiä seisovissa vesistöissä, useinmuodostaa tummanvihreän kalvon veden pinnalle tai mutaiselle pohjalle. Tämän levän solut ovat muodoltaan lieriömäisiä.yhdistetty yhteen säikeeseen.
Nostok - rihmamaiset sinilevät, filamentittai sen ketjut ovat yhteydessä pesäkkeiksi, jotka ovat usein muodoltaan pallomaisia,luumuhedelmän kokoinen. Ulkopuolelta nämä pesäkkeet ovat peitetty hyytelömäisellämassa. Nostok asuu lampien ja järvien rannoilla, kostealla maaperällä.ja säiliöiden pohjalla.
Joitakin sinileviä muodostuu yhdessä sienten kanssaerilaisia jäkälätyyppejä.
Sekanivelsiteiden laitos merilevää
Multiflagellate tai kelta-vihreä,leville on ominaista se, että niiden zoosporeissa on2 epätasaista siimoa, joista lyhyt siima on sileä ja pitkä siima on pinnallinen. Kromatoforit kellanvihreät, kiekon muotoiset.Yksisoluiset, koloniaaliset, rihmamaiset ja ei-soluiset organismit.Esimerkki on botridium.
Botridium- tyypillisinmultiflagellate levien istuttaja. Tallus on halkaisijaltaan 1-2 mm vihreä rakkula rakkulan alapuolellavärittömät haaroittuneet kasvut sijaitsevat - risoidit, jotkalevää on upotettu maaperään. Se on yksisoluinen monitumamerilevää. Vesikkelin keskiosa on täynnä solumehlaa, sytoplasma sijaitsee parietaalisesti. Kromatoforit sisältävät paljon caroatinoideja, joten botridiumilla on kellertävänvihreä väri.Pyrenoidit puuttuvat.
Lisääntyy pääasiassa zoosporien avulla, joita tuotetaanvaltavassa määrässä kuplan keskellä. Elää kosteallamaa, lätäköiden reunoja pitkin, muodostaa tummanvihreitä ryöstöjä maaperäänvärit.
Piilevien laitos merilevää
Piilevät, piilevät tai piidioksidit tai basillit, levät ovat äärimmäisiäteelajike, mikroskooppisesti pieni, enimmäkseenyksisoluisten organismien tapauksista. Tämä leväosasto on ominaistarizovaetsya erikoinen solujen rakenne. Kenno on peitetty kiinteällä aineellakuori pektiinin muodossa, hyytelömäinen kalvo, ulkopuolellasitä peittää piikivikuori, joka koostuu kahdesta itsestäänseisovat puolikkaat, niin sanotut puitteet. Yksi näistäkivi peittää toisen kuin laatikon kansi. Puolikkaita pitkinkuoressa on rakomainen aukko molemmilla puolilla. Sen läpisolun sytoplasman avautuminen kommunikoi ulkoisen ympäristön kanssa. kohdeki on poikkeuksellista voimaa. Ne eivät sulataXia eläimet ja linnut eivät tuhoudu edes hehkuvanatulessa. Lajia on yli 5000. Omaperäiset näkymätPääpiirteet ovat solujen muoto ja erilaiset paksunnuksetkuoret arpien, verkkojen jne. muodossa.
Tietyntyyppisten piilevien solujen muoto onpitkänomainen neliö, elliptinen, pyöreä, tähden muotoinen, muodossanauhat, spiraalit jne. Solut sisältävät sytoplasman, ytimen ja yhdentai useampia kromatoforeja. Klorofyllin ja fukoksantiinin lisäksiKromatoforit sisältävät myös muita kellanruskeita pigmenttejä,siksi kromatoforit ovat keltaisia. Piilevien soluissa ei ole tärkkelystä, vara-aineita edustaapellavaöljy.
Ne lisääntyvät pääasiassa suoraan jakautumalla, mikähe kulkevat omalla tavallaan. Jakamisen aikana jokainen tytärsoluvastaanottaa ytimen, yhden kromatoforin ja vain yhden kuoriventtiileistä,toinen taite muodostetaan uudelleen. Suoran jaon lisäksi diatomerilevät lisääntyvät seksuaalisesti fuusioituessaankaksi solua, jotka ovat aiemmin luopuneet kuorensa.
Piilevät elävät meri- ja makeissa vesissäusein ne ovat pääasiallisia olennainen osa planktonia ja ne ovat arvokasta ravintoa eläimille. Kuolleiden leväsolujen lehdet vajoavat pohjaan ja muodostavat vähitellen valtavia kerrostumia,tunnetaan nimellä "vuorijauho, piimaa, tripoli.
Suuret piileväkertymät piilevien muodossa jala ovat keskittyneet valtamerten napa-alueille, lähellä Alaskaa,Aleuttien saaret Okhotskinmerellä ja Beringinmerellä. Saatavillane ovat Itämerellä.
Piileviä ovat makean veden levä pineulusria,fragilariaja raporttikorttiria; nämä levät muodostavat pesäkkeitä nauhojen muodossatai ketjut;naviculatuuhea kolo kasvaaniei.
Viherlevien laitos
Viherlevä on yksi monipuolisimmista leväryhmistä, se yhdistää noin 5000 lajia.Tämän osaston edustajille on ominaista vihreä väri,joka johtuu klorofyllistä ja jota mikään ei peitätai muita pigmenttejä. Vihreitä leviä edustavat yksisoluiset, monisoluiset, siirtomaamuodot. Vihreätlevillä on useimmiten rihmamainen rakenne, filamentit koostuvatyksi rivi soluja. Solussa on selluloosakalvoplasma, ydin ja kromatoforit. Lisääntyminen tapahtuu aseksuaalisestivegetatiivisesti ja seksuaalisestiviherlevät elävät vedessä,Mutta jotkut heistä asuvat myös maallaaiheita.
Suurin osa esittääiley elääpuissa, kivien päällänyah kosteissa, varjoisissa paikoissa. Lammissa ja joissa muodostuumuta.
Viherlevien osasto on jaettu useisiin luokkiin, joistaTarkastellaanpa yhtäläisiä flagella- eli vihreitä, pariutuvia leviä tai konjugaatteja, ja hiiltä tai säteitäLuokka equiflagellate Vihreän veden joukossakasvaa, tämä luokka on laajin. Edustajia on monenlaisiaulkonäössä ja sisäisessä rakenteessa. Ne ovat ominaisiakahden identtisen siimon läsnäolo Tämä luokka yhdistää yhdensolu- ja siirtomaamuodot, liikkuvat ja liikkumattomat muodot.Luokka on jaettu 8 luokkaan. Tarkastellaan edustajia 4 byrivit.
Tilaus volvox . Suurin osatyypillisiä Volvox-järjestyksen edustajia ovatklamydomonas ja volvox.
Chlamydomonas on yksisoluinenliikkuva merilevä. Asuu suuria määriämakeissa matalissa vesistöissä - lätäköissä, lammissa, ojissateiden varrella. Runsaan lisääntymisen ansiosta se muuttaa veden vihreäksiväri. Se on mikroskooppisesti pieni soikea solutai pyöreä muoto. Toisesta (etu)päästä häkki on pitkänomainennokan muodossa, samassa päässä on 2 samanlaista lippua, jotka edistävät levien liikkumista. Koko solun ontelo on täytettyjoita ei kata sytoplasma, jossa tuma sijaitsee, lähempänä nenää
Volvoks taihyrrä, on tyypillinen esimerkki mikroskooppisten levien siirtomaamuodoista. Tämän levän pesäke näkyy paljaalla silmällä, se saavuttaaneulanpään kokoinen ja pallon muotoinen. Tällainen pallomainen Volvox-pesäke koostuu valtavasta määrästä soluja (jopa 50 tuhatta), jotka sijaitsevat yhdessä kerroksessa pallon reunalla.Jokaisessa solussa on 2 flagellaa. Kaikki flagellat sijaitsevat vieressäja helpottaa koko pesäkkeen liikkumista. LiikeKaikkien solujen flagellat ovat aina koordinoituja. Pallon ontelo on täytettyei ole peitetty nestemäisellä limalla. Volvox lisääntyy vegetatiivisesti jaseksuaalisesti. Vegetatiivisen lisääntymisen aikana äidin sisälläpesäke (pallo) Muodostuu 8 ... 15 tytäryhdyskuntaa (palloa). klotytäryhdyskuntien kypsyminen, aikuisen pallon seinät ovat repeytyneetja nuoria pesäkkeitä tulee ulos, minkä jälkeen emokolonia kuolee.
Seksuaalinen lisääntyminen - oogamia - tapahtuu vain useimmissalee suurempia soluja. Samaan aikaan Volvox-yhdyskunnan soluissa noinpaljaat kaksileipäiset sukusolut erotetaan toisistaan, vain levän eri yksilöiden sukusolut sulautuvat yhteen.
Volvox-tyyppejä on monenlaisia. Kaikki ne ovat yleisiäpääasiassa seisovassa, hyvin lämmitetyssä makeassa vedessävesisäiliöt, joissa on seisovaa vettä - lammet, järvet, harvemmin joissa
Chlorellamikroskooppinen yksisoluinenpallomainen merilevä. Chlorellalle on ominaista nopealisääntyminen ja erittäin aktiivinen fotosynteesiprosessi. Kiitoksetsuuren määrän klorellaplastidien läsnäololla on erilainen vaikutusaktiivista käyttöä aurinkoenergia, perinteinen kulttuurikasvit kuluttavat vain 0,1 % aurinkoenergiasta biokemiallisestiikaaliset muutokset ja chlorella -2,5%, eli 25 kertaa enemmän.muu positiivinen laatu chlorella - erittäin nopea kasvuzhenie.
Ulothrix on rihmamainen, haarautumaton levä,joka koostuu yhdestä solurivistä. Pohja väritön lankahäkkion omituinen pitkänomainen muotoTämän solun avulla levät kiinnittyvät vedenalaisiin esineisiin.
Ulotrix-filamentti pitenee poikittaisen solujakautumisen vuoksija voi kasvaa loputtomiin. Solut ovat homogeenisialyhyt, jokainen niistä sytoplasma, ydin, kromatoformitpyrenoidien kanssa. Ulothrix lisääntyy pääasiassa aseksuaalisesti.muodostamalla 4-leimattuja zoosporeja. Nämä zoosporit ovataika liikkuu ja asettu sitten johonkin vedenalaiseen esineeseenja kasvaa uudeksi säikeeksi.. Seksuaalinen prosessi on isogamiaa. Lisäksi miehen ja naisen sukusolut ovat ulkoisesti erottamattomia, mutta fysiologisialoogisesti ne ovat erilaisia ja tulevat eri säikeistä, niin erilaisiasukusolut on merkitty + ja -.
Ulothrix elää makeassa vedessä, jossa ne kasvavat allavesiesineet (kivet, kasat), jotka muuttuvat kirkkaan vihreiksiväri.
Cladophora on rihmamainen vihreä levä.Tallus koostuu yhden rivin muodostamista haarautuvista langoistamoniytimiset solut, yksittäiset kasvit näyttävät vedenalaisiltapensaat Solut ovat suuria, moniytimiä, paksuja,kerroksellinen kuori. Löytyy makeista ja suolaisista vesistä
Division punainen levää tai tulipunaista naschiaSiellä on yli 600 sukua ja noin 4000 lajia. muista levistäeroavat väriltään, mikä johtuu läsnäolostaklorofyllin lisäksi niissä on kaksi muuta pigmenttiä - fykoerytriini (punainen) ja fykosyaani (sinivihreä). SuhteestaLevien väri riippuu näiden pigmenttien muodostumisesta (se vaihteleepunaisesta melkein mustaan). Osaston edustajat suurimmassa osassa monisoluisiauudet organismit monimutkainen rakenne, ja vain alkeellisimmatheillä on yksisoluinen tai siirtomaa-tallus. Useammin thalluson pensaiden, levyjen muodossa, joissakin se on erittäin voimakkaasti leikattu.
Punalevät elävät merissä suuremmissa syvyyksissä kuinvihreä ja ruskea. Tämä johtuu punaisesta pigmentistäedistää vihreiden ja sinisten säteiden sieppaamista suurissa syvyyksissäspektrin ja siten parantaa fotosynteesiprosessia. Punaisessaleväkertymä ei tavallinen tärkkelys. Toisin kuintavallinen tärkkelys, sitä ei värjätä jodilla sinisellä, vaan punaisellaruskea väri.
Punalevistä puuttuu eläinporit ja siittiöt.Aseksuaalinen lisääntyminen niissä tapahtuu liikkumattomien avullariitaa. Seksuaalinen lisääntyminen - oogamia, mutta siittiöiden sijaanmuodostuu miessoluja - siittiöitä, ne siirtyvät muniin veden avulla. Oosyytit tuotetaan erityisissä elimissä -carpogonit.
_Punalevät sisältävätjälkiruokatalluksella pensaan muodossa; polysifoniajossa talluksella on haarautuva lanka.
osasto ruskea merilevää
Ruskealeväosaston edustajilleoranssin ruskea väri kromatoforien esiintymisen vuoksiruskea pigmentti - fukoksantiini. Fukoksantiinin läsnäolopeittää vihreän värin ja antaa näille leville ruskean värinerilaisia sävyjä. Fukoksantiinin lisäksi ne sisältävät ksantoafilo ja karoteeni. Ruskealeväosasto yhdistyy900 lajia.
Ruskeat levät erottuvat yleensä suuresta monisoluisesta talluksesta. Suurimmat levien edustajat löytyvät juuri ruskealevistä. Jotkut niistä, kuten unikkorocystis, saavuttaa 60 metrin pituuden, mutta on myös pieniä muotojamuutaman millimetrin etäisyydellä toisistaan.
Tärkkelyksen sijasta ruskealeväsolut sisältävät glukoosiaja sokeripitoiset aineet - kutsuvat ja rakkolevä, jotka antavatNäillä levillä on makea maku kypsennettynä. Kutenne keräävät usein öljyjä.
Ruskealevän talli on monivuotinen, mutta lehtimäinenlevyt kuolevat vuosittain ja kasvavat uudelleen aikaisin keväällä.
Ruskealevien monimutkainen ulkoinen rakenne määrääkloniitä ja anatomisen rakenteen erilaistuminen (heillä onerilaisia solumuotoja). Jotkut tutkijat uskovat, että nämälevillä on jopa erilaisia kudoksia.
Ruskeat levät lisääntyvät monin eri tavoin. Nekojotkut heistä lisääntyvät primitiivisillä seksuaalisilla keinoilla - isomia, kun 2 samanmuotoista sukusolua sulautuvat yhteen. muut,kehittyneemmät levät (levä) on monimutkaisempiseksuaalinen prosessi - oogamia, jossa suuri munasulautuu pienen liikkuvan urospuolisen sukusolun - siittiöiden - kanssatalo.
Ruskeissa levissä tapahtuu suvutonta lisääntymistäzoosporit, joita tuotetaan suuria määriä zoospossariveissä. Ruskeissa levissä sukupolvien vuorottelu ilmenee melko selvästi; aseksuaalista ja seksuaalista. Lehtien terien päällänämä levät muodostavat yksisoluisia zoosporangioita, jotka kerätäännye ryhmiin, joiden joukossa ovat hedelmättömät langat.Jokainen zoosporangium tuottaa 16...64 tai enemmäneläinporit. Zoosporit ovat ulkoisesti identtisiä, mutta fysiologisesti erilaisia.Jotkut heistä itävät ja muodostavat mikroskooppisen pieniä vaimoja.skye ja muut - miespuoliset gametofyytit. urospuolisille gametofyyteillemuodostuu lisää anteridia ja niissä yksi siittiözoidi, ja naaraspuolisille gametofyyteille muodostuu oogonia, joka kantaayksi kananmuna. Siittiöiden ja munasolujen yhdistämisen jälkeenmuodostuu tsygootti, josta kehittyy aseksuaalinen sukupolvi -sporofyytti.
Ruskeat levät ovat meren elämää, monet niistäne ovat yleisempiä pohjoisilla merillä, usein niitä muodostuuvaltavia metsiä merissä ja valtamerissä. Atlantin pohjoispuolellavaltameressä, Sargasson meressä, valtavassa määrässäOn olemassa ruskealeväsuku - Sargassum. Nämä levät ovat enemmänkaikista ovat kelluvassa tilassa läsnäolon vuoksierityisiä ilmalla täytettyjä kuplia.
Ruskealeviä pidetään muinaisena kasviryhmänäkorkeampi erilaistumisaste löytyy paitsiulompi, mutta myös talluksen sisäosa. Ulkoisesti ne ovat samanlaisiashimi-kasveja, joten jotkut kasvitieteilijät uskovat, että nämälevät ovat saattaneet synnyttää korkeampia kasveja.
Ruskealevädivisioona koostuu 4 lajikkeesta. Harkitse ennenkahden luokan putterit: rakkolevä ja fucus.
Tilaa rakkolevä. Tämäerittäin suuret levät, joskus jopa 60 metriä korkeat.Heidän talluksensa on voimakkaasti leikattu ja lisäksi hyvin kehittynythaarautuneet risoidit, joilla levät ovat tiukasti kiinni meren pohjassa. Laminariat elävät merten rannikkokaistaleella5 ... 10 metrin syvyydessä ja muodostavat usein valtavia vedenalaisia paksuja"metsät".
Laminaareihin kuuluvat suku Laminaria (sisältää 30 lajia), suvu Lessonia (sisältää5 lajit) ja Macrocystis-suku. Nämä levät ovat monivuotisia kasveja, jotka eroavat toisistaantalluksen rakenne.
osasto lima muotit
Plasmodium muodostuu paljaiden ameevien fuusiossalimamuotin eri soluja ja joissakin limamuottityypeissä ulottuuihmisen kämmenen kokoa.Hän yleensä väriltään kirkkaan keltainenväri ja sillä on kyky olla hyvin hitaasti amebamainenliikkua (0,1 mm/min). Liikkuessaan plasmodiumilla on taipumuspiiloudu valolta ja suuntaa kohti kosteuden lähdettä. Tavataliman homeita löytyy yleensä varjoisista metsistä; mätäneillä kasveilla, välilläkuori ja puu, halkeilevissa kannoissa, pudonneiden lehtien alla.Ne lisääntyvät itiöillä.
Limamuottien sijainti fylogeneettisessä järjestelmässä on epäselvä,ilmeisesti ne polveutuvat jonkinlaisesta siimosta. Tästäosastolla, harkitse plasmodioforia.
Tauti leviää maaperässä ja kehittyy erityisen nopeasti.viihtyy happamassa maaperässä.
Kaikilla heterokonteilla näyttää olevan flagellaarinen laite. Siimaloita on 2, ja toisessa niistä on erittäin tyypillisiä putkimaisia kolmijäsenisiä siipiä, eli karvoja - mastigonemeja. Kahden epätasa-arvoisen siimon läsnäolo oli perustana tälle laajalle vesieliöryhmälle. monilipuinenlevät, tai heterokontami. Pituuserojen lisäksi siima eroaa myös morfologisesti: pääsiima koostuu akselista ja sen päällä olevista värekarvoista, sivusiima on sileä ja siimamainen. Pigmenttien koostumus on myös samanlainen - klorofyllit A Ja Kanssa ja lukuisia karotenoideja sekä kloroplastien rakennetta. Kloroplasteja ympäröi neljä kalvoa, joista kaksi ulkokalvoa on liitetty solun endoplasmiseen retikulumiin.
Osana Heterocont (Diversified) leväosastoa useita leväryhmiä on yhdistetty luokkiin: Brown, Golden, Yellow-Green, Rafidophytes, Sinura, Feotamnia, Eustigma, joita pidettiin aiemmin itsenäisinä osastoina.
Luokka Ruskealevä– Phaeophyceae (Fucophyceae)
Luokan nimi "ruskealevä" heijastaa niiden talluksen väriä (kreikasta. Rhaios- tumma). Väri johtuu suuresta määrästä ruskeita ja keltaisia pigmenttejä - klorofyllien lisäksi A Ja Kanssa Ja β -karoteeni niiden kloroplasteissa on ylimäärä ksantofyllejä, erityisesti fukoksantiinia.
Monisoluiset, pääasiassa suuret levät, erityisen laajalle levinneitä molempien pallonpuoliskojen kylmissä vesissä. Ruskealevistä ei tunneta yksisoluisia eikä siirtomaamuotoja. Noin 1,8 tuhatta lajia tunnetaan.
Solun rakenne. Ruskealevälajin solu on eukaryoottinen: se on muodostanut organelleja, joissa on omat kalvonsa: ydin, mitokondriot, plastidit jne. Vihreät pigmentit - klorofyllit A Ja Kanssa- peitetty lisäaineilla: oranssit karoteenit ja kellanruskeat ksantofyllit; jälkimmäisistä fukoksantiini on hallitseva. Ruskealevien soluista löytyy plastideja yhdestä useaan. Ulomman solukalvon päälle ruskealeväsolut kehittävät yleensä jäykän soluseinän.
Ydin on useimmiten yksi, mutta joskus ytimiä on useita. Joillakin edustajilla ytimien lukumäärä kasvaa solun iän kasvaessa. Ytimessä on aina yksi ydin.
Ruskealevien vegetatiivisten solujen kloroplasteissa on silmä (stigma) ja pyrenoidit, joita voi olla yksi tai useampi; joistakin lajeista puuttuu pyrenoidit. 30–40 lipidipallosta koostuva silmäsolu toimii linssinä, joka kohdistaa valon siimaturvotukseen, joka on fotoreseptori. Ruskeiden levien eläinporeilla ja sukusoluilla ei yleensä ole silmää. Vara-aineet kerrostuvat pyrenoideihin. Kloroplastit ovat useimmiten pieniä, levymäisiä, parietaalisia. Niiden muoto voi olla tähtimainen, nauhamainen tai lamellimainen. Ruskealeväsolujen kloroplastien muoto voi muuttua solun ikääntyessä. Kloroplastivaippa koostuu neljästä kalvosta, joista kaksi ulkokalvoa on kytketty EPS:ään, solun endoplasmiseen retikulumiin.
Ruskealevien soluseinän muodostava selluloosa - jota joskus kutsutaan algulloosiksi - eroaa ominaisuuksiltaan korkeampien kasvien selluloosasta. Selluloosa seinämän ulkopuolella on erittäin limainen pektiinikerros, joka muodostuu pääasiassa algiinihaposta ja sen suoloista ja proteiiniyhdisteistä. Algiinihappo löytyy myös vain ruskeista levistä.
Yleiset luonteenpiirteet.
Kaikki ruskeat levät ovat fototrofit. Fotosynteesituotteet - spesifinen hiilihydraatti krysolaminariini,alkoholi mannitoli Ja lipidit. Lipidit kerääntyvät öljypisaroiden muodossa. Krysolaminariini kerrostuu kloroplastin ulkopuolelle: se sijaitsee erityisissä tyhjiöissä sytoplasmassa lähellä pyrenoidia. Mannitoli suorittaa reservin sekä osmoregulatorisen toiminnon; sen pitoisuus solun sisällä riippuu veden suolapitoisuuden muutoksista.
Ruskealevien nuoret solut sisältävät pieniä ja lukuisia tyhjiöitä, jotka kasvavat iän myötä fuusion seurauksena. Sytoplasmassa on erityisiä muodostumia - fysodeja, jotka sisältävät florotaniinit. Nämä aineet tunnetaan vain ruskealevälajeissa. Florotaniinien toiminta voi liittyä leväthallin suojaamiseen eläinten, kuten nilviäisten, syömiltä. Ruskeiden levien florotaniinit todennäköisesti estävät epifyyttisten levien ja eläinten asettumista talluksen pinnalle. Florotaniinien oletetaan osallistuvan säteilyvaurioilta suojaamiseen ja raskasmetalli-ionien selektiiviseen absorptioon vesipitoisesta väliaineesta. Värittömät florotaniinit hapettuvat ilmassa pigmentiksi fykofeiiniksi, joka antaa kuivatuille ruskeille leville niiden tyypillisen tumman värin.
Talluksen rakenne. Ruskealevälajeissa ei ole yksisoluisia ja siirtomaamuotoja, kaikki yksilöt ovat monisoluisia. Ruskeiden levien tallin muoto on monipuolisin. Nämä ovat langat, hiipivät tai pystysuorassa seisovat, yksiriviset tai moniriviset; massiiviset kiinteät tai kaakeloidut kuoret ja kuoret; ohuet limakalvot tai tiheät nahkaiset levyt varressa, yksinkertaiset tai leikatut; kuplat ja pussit; putket ja johdot; vaihtelevasti haarautuneita ohuita pensaita tai voimakkaita pensaita. Thallin kiinnitys suoritetaan risoidien tai pohjien avulla. Pitämään ne pystyasennossa suuret pohjaeläimet muodostavat ilmakuplia, jotka ovat täynnä kaasua (kuva 29).
Riisi. 29. Ruskealevien ulkonäkö: 1 - Fucus; 2 - sakkariini; 3 - Alaria; 4 - Sargassum
Laminaria- ja Fucus-lahkojen edustajien tallit ovat ruskealevälajeista monimutkaisimmat. Niissä on merkkejä kudosten erilaistumisesta tallussolujen erikoistumisesta (kuva 30). Heidän thallissaan voidaan erottaa haukkua joka koostuu voimakkaasti värjäytyneistä soluista ja ydin, joka koostuu värittömistä soluista, jotka kerätään usein filamenteiksi. Laminariassa ytimeen muodostuu myös seulaputkia ja putkimaisia filamentteja. Ydin ei suorita vain kuljetustoimintoa, vaan myös mekaanista, koska se sisältää kierteitä, joissa on paksut pituussuuntaiset seinämät. Monien ruskealevien kuoren ja ytimen välissä voi olla suurien värittömien solujen välikerros.
Korkeus ruskealevien edustajien talli on useimmiten interkalaarinen (intercalary) ja apikaalinen (apikaalinen), harvoin basaali. Interkalaarinen kasvu voi olla diffuusia tai siinä voi olla kasvuvyöhyke. Suurissa muodoissa interkalaarinen (interkalaarinen) meristeemi tai aktiivisesti jakautuva soluryhmä sijaitsee kohdassa, jossa "lehtilehti" siirtyy "lehti" -terään. Suurilla levillä on myös meristaattinen vyöhyke talluksen pinnalla, niin kutsuttu meristodermi (eräänlainen korkeampien kasvien kambiumin analogi).
jäljentäminen ruskealevissä, kasvullisissa, aseksuaalisissa ja seksuaalisissa. Sukusolut muodostuvat monisoluisissa gametangioissa, zoosporit - sisään 
Riisi. 30. Sisäinen rakenne Laminaria(mukaan: L.L. Velikanov et al., 1991): A- varren pituusleikkaus; B- varren poikkileikkaus; SISÄÄN- pala lautasesta, jossa on zoosporangiaa
yksisilmäiset itiöt.
Vegetatiivinen lisääntyminen useissa ruskealevälajeissa voidaan suorittaa talluspaikoilla; lajeissa kiltti Sphacelaria- sikiön oksat (pesimäsilmut); suvun lajeissa Fucus- pohjassa oleva soluryhmä, joka pystyy erilaistumaan uudeksi tallukseksi.
Ruskealevien epäseksuaalinen lisääntyminen tapahtuu yleensä zoosporien avulla. Jotkut lajit lisääntyvät liikkumattomilla tetrasporeilla tai monosporeilla. Aseksuaalisen lisääntymisen itiöt muodostuvat meioosin ja sitä seuraavien mitoosien seurauksena yksisilmäiset itiöt. Seksuaalinen prosessi on isogamia, heterogamia ja oogamia. Sukusolut muodostuvat sisään monitahoinennämä gametangia- erityiset soluryhmät. Jokaiseen gametangiumin "pesään" muodostuu yksi sukusolu.
Ruskealevälle, sukupuoliferomonit- liukoiset aineet, jotka koordinoivat solujen toimintaa lisääntymisen aikana. Feromonit, joista tunnetaan vähintään kymmenen, voivat joko stimuloida anteridiajen avautumista tai houkutella urospuolisia sukusoluja naispuolisiin sukusoluihin. He ovat myös mukana lajien eristämisessä. Uskotaan, että feromoneja muodostuu useimmissa ruskealevälajeissa, joilla on seksuaalinen prosessi.
Elinkaari. Ruskeilla levillä on kaksi päätyyppiä elinkaaret. Yksi tyyppi on haplodiplobiont. Sporofyytit ovat diploideja, ts. Kaikki solut sisältävät täydellisen sarjan kromosomeja. Suvuttoman lisääntymisen itiöt muodostuvat yksisilmäisissä itiöissä. Itiöiden muodostumisen aikana tapahtuu pelkistysjako - meioosi. Tällaista pelkistystä kutsutaan itiöiseksi. Meioosin jälkeen muodostuneet solut (itiöt) ovat haploideja; sisältää puolet kromosomien määrästä. Haploidiset zoosporit ja tetrasporit itävät haploidiksi gametofyytiksi, jolle muodostuu sukusoluja monisoluisissa gametangioissa. Sukusolujen fuusion jälkeen muodostunut diploidinen tsygootti kasvaa diploidiseksi sporofyyttiksi.
Toinen tyyppi on diplobiont; sporofyytit, itiöt ja gametofyytit ovat diploideja; väheneminen jakautuu sukusolujen muodostumisen aikana. Tällaista vähennystä kutsutaan gameettiseksi. Haploidisten sukusolujen fuusion jälkeen muodostuu diploidinen tsygootti, joka itää diploidiseksi sporofyytiksi. Tämä tyyppi on yleinen rakkolevälajeissa (kuva 31).

Riisi. 31. Elinkaari Sakariinit(mukaan: L.L. Velikanov et al., 1991): 1 – sporofyytti, 2 – zoosporangian sori, 3 – eläinporofyytti , 4 - gametofyytti, 5 - anteridium, 6 - oogonium, 7 - anterotsoidi, 8 - tsygootti (hedelmöitetty muna)
Makroskooppisella sporofyytillä Sakariinit, levyn yläosaan kehittyy raidat sori(ryhmät) zoosporangia. Zoosporangioissa muodostuu pelkistysjakautumisen jälkeen zoosporeja, joissa on kaksi eriarvoista siimoa. Ne kasvavat mikroskooppisiksi gametofyyteiksi: uroksilla on anteridia ja naaraalla oogonia. Anterozoidien hedelmöittynyt munasolu itää välittömästi, ilman lepoaikaa ja muodostaa sporofyytin. Tällä elämänkaarella Sakariinit päättyy.
Fucus-levissä ei ole sporofyyttejä. klo Fukusa miehen ja naisen sukuelimet - anteridia siittiöineen ja konseptakkelit, joissa on oogonia ja munat - sijaitsevat talluksen syvennyksissä - scapidit, uros- ja naaraskirvat voivat olla samalla päällä astia tai eri resepteissä. Oogonia istuu häkkijalassa , ilmestyy seinälle käsite(Kuva 32, C, D). Ogoniumsolu jakautuu kolmesti

Riisi. 32. Jäljentäminen Fukusa(mukaan: B. Fott, 1971; W. Braune et ai., 1974): A – talluksen pala (1 – ilmakupla, 2 - resepti); B– viilto astioiden läpi; SISÄÄN– osa naaraspuolisesta scafidiasta; G- oogonia; D- munien vapauttaminen; E– uroskirkkarin leikkaus; JA– haara anteridia; 3 – siittiöiden tuotanto
ytimiä, joista ensimmäinen on pelkistys, jonka seurauksena oogoniumiin muodostuu 8 munaa.
Antheridia muodostuu yleensä haarautuneisiin filamentteihin, jotka tukevat niitä - parafyysejä(Kuva 32, E,JA). Useiden jakautumisten seurauksena muodostuu 64 siittiötä. siittiöt Fukusa silmä sijaitsee pelkistyneen kloroplastin sisällä. Siittiösolun etupäähän muodostuu 13 mikrotubulusta kulkurinykyinen- erityinen rakenne, jonka oletetaan osallistuvan naispuolisen seksuaalisen houkuttimen määrittelyyn.
Siittiöt houkuttelevat munaa Fukusa fucoserraten, munan erittämä epäspesifinen feromoni. Lajitason tunnistaminen liittyy tiettyihin oligosakkarideihin munassa ja siittiössä. Munan pinnalla on erilaisia alueita, joista jokainen sisältää erityisiä orgaanisia yhdisteitä - glykoproteiineja. Siittiöt sisältävät glykoproteiineja, jotka on organisoitunut paikkoihin etusiiman kalvolla ja kasvaimilla sekä itse siittiön pinnalla. Kohdatessaan munasolun siittiö liikkuu munankuoren yli, ikään kuin tutkii sitä etummaisen siimansa avulla. Jos siittiön glykoproteiinit täydentävät munasolun glykoproteiineja, vain munasolu tunnistaa siittiön.
Systematiikka
Keski-Silurialta (noin 425 Ma) tunnetaan fossiilisia löytöjä, jotka voivat liittyä ruskealevään. Mutta nämä löydöt muistuttavat myös joitain nykyajan viher- ja punaleviä. Fossiililöydöt, jotka voidaan ehdottomasti yhdistää nykyaikaisiin ruskealevisiin, ovat peräisin mioseenista (5–25 miljoonaa vuotta). Nämä levät muistuttaa nykyaikaista Dictiotaa. Molekyylimenetelmin määritetty ruskealevien ikä on 155–200 miljoonaa vuotta.
Ruskealeväluokkien erottamiseen käytetään talluksen organisaatiotyyppiä, pyrenoidin läsnäoloa tai puuttumista, kasvutapaa, sukupuolisen lisääntymisen tyyppiä (isogamia, heterogamia, oogamia) ja elinkaarta. Viime vuosina ruskealeväjärjestelmää on tarkistettu aktiivisesti. Eri luokittelujärjestelmissä ruskealevän luokan osana erotetaan 7-14 luokkaa.
Desmarestilainen järjestys–Desmarestiales. Levät ovat tuuheaisia, haaroittuneita, ja niissä on terete, litteä tai litteä tall. Kiinnittyy alustaan hyvin kehittyneellä pohjalla. Talluksen pinnalle kehittyy karvoja.
Suku Desmarestia (kuva 33). Ruskeat tai oliivipensaat, joilla on hyvin erottuva pääverso, jopa 0,8–1 m korkea. Kiinnitetty kartiomaisella pohjalla. Haaroittuvat 3-4 tilausta, vuorotellen tai vastakkain. Viimeisten luokkien oksat ovat litteitä tai harvemmin lankamuotoisia. Lajit ovat merellisiä, elävät arktisilla ja lauhkeilla leveysasteilla 1–20 metrin syvyydessä, muodostaen pensaikkoja tai kasvavat yksittäin.

Riisi. 33. Desmarestia (r. Desmarestia)
Ralphsian järjestys–Ralfsiales. Järjestön edustajilla on tallus eri rakenteiden kuorien muodossa tai ne koostuvat tyvikuoresta ja pystysuorasta versosta.
Suku Analipus. Tallus koostuu monivuotisesta kuorimaisesta tyviosasta ja siitä ulottuvista yksivuotisista, oliivinruskeista, korkeintaan 40 cm korkeista pystysuoraista versoista Monisoluiset itiöt ovat muodoltaan lieriömäisiä ja rakenteeltaan kaksiriviä. Yksisilmäiset itiöt ovat soikeita ja kehittyvät assimilaatiohaarojen joukossa. Suvun lajit ovat laajalle levinneitä lauhkeiden leveysasteiden merillä muodostaen vöitä vuorovesivyöhykkeellä.
Tilaa Dictyota–Dictyotales.Laukun lajeille on ominaista apikaalinen kasvu. Vegetatiiviset solut sisältävät lukuisia kiekon muotoisia kloroplasteja ilman pyrenoideja. Parenkymaalinen tallus, isomorfinen sukupolvien vuorottelu on ominaista. Sukupuoliprosessi on oogaminen, suvuton lisääntyminen liikkumattomilla itiöillä, siittiöissä on vain yksi siima. Siima on pitkä, siipikarvainen ja eteenpäin suunnattu. Luokan edustajia löytyy pääasiassa trooppisista ja harvemmin lauhkeista meristä.
Suku Dictyota (liite, 4A). Sillä on litteä, oliivinruskea, kaksijakoisesti haarautunut, jopa 11–15 cm korkuinen pensas, jonka oksat ovat suoraviivaisia, kapeita, suonittomia, latvahaarukkaisia. Tetrasporangiat muodostavat talluksen pinnalle soreja, joilla on epämääräinen tai rengasmainen muoto. Levitetty lauhkeiden ja lämpimien leveysasteiden merivesissä.
Tilaa Laminaria–Laminariales. Laminarialla on tallus, joka koostuu levystä, varresta ja kiinnityselimistä - pohjasta tai risoideista. Levy on tasainen tai ryppyinen, kokonainen tai leikattu, reikiä, poimuja, petsejä tai ilman niitä, useista kymmenistä senttimetreistä 20–40 m pitkä Varsi on haarautunut tai useammin haarautumaton. Sporofyytit ovat yksivuotisia tai monivuotisia, jotkut lajit ovat 11–18-vuotiaita. Limakanavat sijaitsevat monien rakkolevän kuoressa.
Ruskealevien suosituimmat edustajat ovat rakkolevä (sukutyypit Laminaria tai Saccharina), tai "merilevää", joita levitetään pääasiassa Tyynellämerellä. Niitä löytyy veden pinnasta (reunasta) 200 m syvyyteen asti. Laminaria kasvaa mieluiten 2–20 (30) m syvyydessä, paikoissa, joissa vesi liikkuu jatkuvasti, kiinnittyen kiviin ja kiviin. Laminariat muodostavat levävyöhykkeitä Kaukoidän merien rannikolla. Voimakkaimmat pensaat löytyvät Kurilien saarilta ja noin klo. Hokkaido, jonne noin puolet maailman laminariavaroista on keskittynyt. Maailman rakkoleväsato saavuttaa 2 miljoonaa tonnia tuorepainoa, kun taas yli miljoona tonnia tulee sen meriviljelyn tuotannosta Kiinassa.
Suku Sakariini (Laminaria). Tallus on 0,5–5–10 m pitkä, siinä on paksu kiiltävä levy, elastinen varsi ja tiiviisti toisiinsa kietouttuneet risoidit, jotka on kiinnitetty erilaisiin kiinteisiin alustoihin. Levyn reunat ovat sileät tai aaltoilevat, levyn pinnassa on usein pullistumia ja taitoksia. Levyjen molemmille puolille muodostuu itiöpesäkkeitä - sori -, itiöiden vapautumisen jälkeen levyn yläosa tuhoutuu. Elää kahdesta useaan vuoteen. Sakariinit ovat arvokas teollisuuden raaka-aine ja meriviljelyn kohde. Saccharina japanilainen muodostaa pensaikkoja Japaninmeren pohjoisosassa, samoin kuin lähellä Sahalinin etelä- ja kaakkoisrannikkoa ja Etelä-Kuriilisaaria. Saccharina japanilainen on arvokkain kaupallinen laji, sillä on erinomainen maku ja sitä käytetään erilaisten ruokien, säilykkeiden ja makeisten valmistukseen.
Tilaa Sphacelariae–Sphacelariales.
Tähän lahkoon kuuluvilla ruskeilla levillä on haarautuva talli, jossa on apikaalista kasvua. Karvaiset oksat. Vegetatiivisissa soluissa on lukuisia kloroplasteja ilman pyrenoideja. Enemmistön elinkaaren aikana isomorfinen sukupolvien vaihto, seksuaalinen prosessi on isogamia tai heterogamia. Levät muodostavat pieniä, enintään 3 cm korkeita ruskeita rypäleterttuja ja jopa 20 cm korkeita haarautuneita pensaita kiville tai muille leville; on endofyyttisiä edustajia.
Suku Sphacelaria. Sillä on ohut tallus. Haaroittuvat filamentit muodostavat pieniä, jopa 0,5–3 cm korkeita, ruskeita kimppuja. Vegetatiivisen lisääntymisen suorittavat erikoistuneet, usein kolmipalkiset sikiön oksat - leviävät. Seksuaalinen prosessi on isogaminen. Naisten sukusolut erittävät feromoneja ectocarpen ja multifiden. Suvun lajeja on sekä polaarisilla että trooppisilla leveysasteilla.
Tilaa Scytosiphon–Scytosiphonales. Edustajat, joilla on suolisto-, rakkulaarinen, lamellimainen tai sakkulaarinen, usein ontto talli, jonka pituus on enintään 25 cm. Kuori on yksirivinen tai kaksirivinen, talluksen sisäosa (seinä) muodostuu useista kerroksista suuria soluja. Talluksen pinnalle kehittyy monisoluisia itiöitä, jotka kerääntyvät soriin, jotka tukevat niiden lankoja - parafyysejä ja hiuskimppuja.
Suku Scytosiphon on ohutseinäinen ruskea 4–25 cm pitkä putki (Liite 4B). Tallus on kiinnitetty pienellä pohjalla ja siinä on harvinaisia poikittaisia supistuksia. Sitä esiintyy merien rannikkovyöhykkeellä eri leveysasteilla, muodostaen usein vyöhykkeitä vuorovesivyöhykkeellä.
Fucus tilaus–Fucales. Heillä on monivuotinen talli suurten, epäsäännöllisesti tai kaksijakoisesti haarautuvien, jopa 1–3 metrin pituisten pensaiden muodossa. Ne on kiinnitetty alustaan kartiomaisella pohjalla. Talluksessa on versoja, rypäleitä (lehtimäisiä muodostelmia), säiliöitä (sukusolujen säiliöitä) ja uivia (ilma) rakkoja. Niitä löytyy kaikkien lauhkean ja arktisten merien vuorovesivyöhykkeeltä, missä ne muodostavat voimakkaita paksuja - vöitä.
Suku sargassum(Kuva 34, SISÄÄN). Runsaan haarautuneet oliivinruskeat tai ruskeat pensaat, korkeus 0,5–1,5 m, kiinnitetty kartiomaisella pohjalla. Fylloidit kehittyvät sivuversoissa, jotka ulottuvat lieriömäisestä varresta talluksen ala- ja keskiosissa. Phylloidit ovat nahkamaisia, keskikylkeä, 0,2–2 cm pitkiä. Uimarakot ovat soikeita tai pyöristettyjä, kasvuston kanssa tai ilman. Sargassumit ovat laajalle levinneitä lauhkeilla ja subtrooppisilla leveysasteilla alemmasta rannikosta 10 metrin syvyyteen, missä ne muodostavat tiheitä tai harvoja metsikköjä. Eteläisellä Tyynellämerellä - niin kutsutulla Sargassomerellä - ne muodostavat valtavia kelluvia kasaumia, joissa ne lisääntyvät vain kasvullisesti.
Suku Fucus (kuva 32).Thallus nahkainen, tuuhea, oksat kaksijakoisesti, 25–50 cm korkeat.Lähellä Kaukoidän merten rannikkoa, kasvaa useita vuosia, elää rantavyöhykkeellä. Muodostaa usein suuria paksuja matalassa vedessä. Tämän suvun lajeille on ominaista kaksijakoisuus

Riisi. 34. Fucus-levien ulkonäkö: A – Ascophyllum; B – Cystoseira; SISÄÄN – sargassum; 1 - astia, 2 - uimarakko
haarautunut tallus litteillä oksilla, jossa on pitkittäinen kylkiluo, joka on kiinnitetty kiviin kartiomaisella pohjalla. Fucuseja käytetään lannoitteina, karjan rehuna, rehujauhon, alginaattien valmistukseen, käytetään lääketieteessä struuman hoitoon, painonkorjaukseen jne.
Tilaa Ectocarps–Ectocarpales. Tähän sisältyvät levät ovat mikroskooppisia, vapaasti eläviä, epifyyttejä, epitsoiitteja tai endofyyttejä. Sukuelimiä edustavat yksisilmäiset (sporangiat) ja monisilmäiset (gametangia) itiöt; ne ovat pitkiä lieriömäisiä, lyhytsylinterimäisiä tai tyynyn muotoisia.
Suku ectocarpus. Tallus pehmeiden ruskeiden pensaiden muodossa, korkeintaan 2 cm. Holkit muodostuvat yksirivisistä, vuorotellen, kaksijakoisista tai väärin haarautuneista lankoista. Monisoluiset itiöt ovat palomaisia, muodostuneet sivuhaarojen kylkeen 1–3-soluiseen varteen. Lajeja esiintyy eri leveysasteilla olevissa merissä, puhtaissa ja saastuneissa vesissä, rannikolla ja sublitoraalin yläosassa. Epifyyttejä löytyy myös ihmisperäisten substraattien likaantumisesta.
Ekologia
Suurin osa ruskealevistä elää merivesissä. Vain 8 lajia tavataan makeissa vesissä. Siellä on sekä yksivuotisia että monivuotisia lajeja, joiden ikä voi olla 15–18 vuotta. Laminaria levä Nereocystis Ja Makrocystis, jotka elävät Pohjois-Amerikan Tyynenmeren rannikolla, ovat kaikkien levien enimmäiskoko - jopa 30–40 m pitkiä. Ne muodostavat jättimäisiä vedenalaisia metsiä Kaukoidän merissä. Ruskeat levät kiinnittyvät monenlaisiin substraatteihin - kiviin, kiviin, soraan, merieläinten kuoriin ja kuoriin sekä muihin leviin. Jotkut ruskealevän pienet muodot elävät muiden levien kudoksissa endofyytteinä. Useimmat ruskeat lajit elävät kiinnittyneessä tilassa. Maasta irti repeytyneet tallit kulkeutuvat virran mukana tyynille mutapohjaisille paikoille ja ovat siellä edelleen olemassa. Lajit, joilla on ilmakuplia talluksessa, irtautuvat maasta, kelluvat pintaan muodostaen suuria kelluvia klustereita erityisesti alueilla, joilla on vakaa ympyrävirtaus, kuten Sargassomerellä. Ne esiintyvät tällaisissa klustereissa melko pitkään ja lisääntyvät vain kasvullisesti.
Ruskealevät ovat laajalle levinneitä kaikissa merissä, mutta ne saavuttavat suurimman kehittymisensä lauhkeiden ja alipolaaristen leveysasteilla. Niiden suuret tallit kasvavat kiinnittyneenä kallioihin ja kiviin, ja hiljaisissa paikoissa lähellä rannikkoa ja suurissa syvyyksissä niitä voidaan pitää jopa nilviäisten kuorien ja soran venttiileissä. Voit tavata niitä eri syvyyksillä - rantavyöhykkeeltä, jossa laskuveden aikaan ne ovat poissa vedestä tuntikausia, jopa 40–200 m. Havaijin saarten alueella 180 metrin syvyydessä lajit suvusta sargassum, A Rakkolevä V Adrianmeri löytyi 200 m:n syvyydestä. Runsaat ruskealeväpeikot havaitaan 6–15 m:n syvyydessä, missä ovat parhaat valaistusolosuhteet ja jatkuva veden liike, joka tuo ravinteita niiden talliin ja rajoihin. kasvissyöjien asuttaminen - fytofagit.
Luokka kultalevät -Chrysophyceae
Yksisoluiset, siirtomaa- tai monisoluiset organismit, joiden elinkaaressa on yleensä siimavaihe. Solut ovat yksitumaisia, ja niissä on yksi tai useampi kloroplasti. Stigmaa esiintyy yleensä (kuva 36). Flagella 1–2, eriarvoinen. Supistumisvakuoli sijaitsee solun etuosassa. Näiden organismien solujen ulkokuoret ovat hyvin erilaisia. Primitiivisimmissä lajeissa solu on pukeutunut herkälle periplastille, joka mahdollistaa erimuotoisten ulkonemien muodostumisen (rhizopodia, pseudopodia), toisissa plasmalemma on peitetty kovalla selluloosakalvolla, joskus voimakkaasti limaisella. Kultalevissä olevia taloja on eri muotoisia: maljakon muotoisia, pallomaisia, munamaisia, lieriömäisiä; yhdellä tai useammalla reiällä. Häkki kiinnitetään talon pohjaan joustavalla jalalla tai voi olla vapaa. Joissakin krysofyyteissä solun pinnalle kertyy erimuotoisia ja -kokoisia kalkkipitoisia muodostelmia - kokkoliitteja, ja piisiipillä on sisäinen piirunko. Näiden luurankojen rakenteen monimuotoisuus mahdollistaa fossiilisten piiflagellaattien eri muotojen jäänteiden käyttämisen niitä sisältävien geologisten kivien iän määrittämiseen. Kehittyneimmillä kultaleväosaston edustajilla solut on puettu kuoreen, joka koostuu piisuomuista (joskus on piikkejä) tai suljettu taloihin, joiden aukkojen kautta tulee ulos lippuja tai pseudopodia.
Kultalevälajeihin kuuluvat talot ovat maljakon muotoisia, pallomaisia, munamaisia, lieriömäisiä, ja niissä on yksi tai useampi reikä. Häkki kiinnitetään talon pohjaan joustavalla jalalla tai voi olla vapaa.
Pigmentit - klorofyllit A Ja Kanssa, fukoksantiini. Krysofyyteille on ominaista korkein fukoksantiinipitoisuus heterokonteista (55-93 % kaikista karotenoideista).
Suurin osa lajeista on makean veden fototrofeja, mutta myös heterotrofeja ja fagotrofeja löytyy.
Vara-aineet - krysolaminariini ja öljyt.
Kultalevien yksisoluisista muodoista vallitsevat makean veden planktonlajit ja monisoluisista pohja- tai epifyyttiset lajit. Niitä löytyy pääasiassa viileistä merestä ja makeista vesistöistä. Noin 360 lajia tunnetaan.

Riisi. 36. Kultalevät (C. Hoek van den et al., 1995 mukaan): A- solun rakenne; B- radikulaarinen järjestelmä; SISÄÄN- kolmiosainen mastigonema; G- kloroplastin osan leikkaus; 1 - perusrunko; 2 - tyviturvotus; 3 - lyhyt siima; 4 - stigma; 5 - risoplasti; 6 - ydin; 7 - ydinkalvo; 8 - ydin; 9 - mitokondrio; 10 - vakuolit; 11 - lima; 12 - plasmalemma; 13 - rakkulat; 14 - limakalvo; 15 - lipidit; 16 - kloroplasti; 17 - Golgi-laitteet; 18 - supistuva vakuoli; 19 - pitkä flagellum: 20 - 1. juuri; 21 - 2. selkäranka; 22 - 3. selkäranka; 23 - 4. juuri; 24 - kloroplastinukleoidi; 25 - HES; 26 - kloroplastikuori; 27 - vyön lamelli; 28 - lamelli; 29 - pitkä lateraalinen filamentti; 30 - lyhyt lateraalinen filamentti; 31 - päätelanka; 32 - mastigoneeman putkimainen osa
talluksen rakenne kultalevissä se on useimmiten monadinen, mutta voi olla hyvin monimuotoinen: kokkoidinen, palmelloidinen, plasmoidinen, ameboidinen, rihmamainen, parenkymaalinen (kuva 37).

Riisi. 37. Kultalevät (L.L. Velikanov et al., 1981 mukaan): A – Ochromonas: 1 - ulkonäkö, 2 - kysta; B – Chromulina: 1 - ulkonäkö, 2 - kysta, 3 - kaavio kalvon muodostumisesta kystasta; SISÄÄN – Dinobrion: 1 - yleinen näkymä pesäkkeestä, 2 - kysta; G – Chrysameba; D – gibberdia; E – Hydrurus: 1 - yleiskuva pesäkkeestä, 2 - haaran kärki, 3 - zoospore
jäljentäminen. Kultalevissä on kolme lisääntymismenetelmää: vegetatiivinen, aseksuaalinen ja seksuaalinen.
Vegetatiivinen lisääntyminen tapahtuu pitkittäisen solunjakautumisen tai pesäkkeen hajoamisen osiin seurauksena solun jakautumisesta yhteen, kahteen tai kolmeen suuntaan.
seksuaalinen prosessi(kultalevien solut ovat diploideja) - isogamia tai konjugaatio. Seksuaalisen prosessin seurauksena sekä epäsuotuisissa olosuhteissa muodostuu kystat, joissa on paksu piitä sisältävä kuori.
suvuton lisääntyminen yksi- tai kahdella leimautuneella zoosporilla, jotka kehittyvät tavallisissa vegetatiivisissa soluissa tai zoosporangioissa. Seksuaalinen prosessi: holo-, iso- ja autogamia. Hologamia on eräänlainen seksuaalinen prosessi, jossa kaksi vegetatiivisten solujen kaltaista solua sulautuvat etupäistä kaksitumaiseksi tsygootiksi, joka sitten muuttuu silikoituneeksi kystaksi.
Systematiikka
Kultalevistä eristettiin viime vuosisadan lopulla molekyyligeneettisten tutkimusten tulosten perusteella luokka Sinuraceae. Luokka Feotamniaceae erotetaan myös - keltavihreistä ja kultalevistä. Siksi Chrysophyceae-luokan soveltamisalaa on supistettu ja se sisältää tällä hetkellä noin 360 lajia, jotka on luokiteltu kolmeen luokkaan: Chromulinales, Hibberdiales ja Hydrurales. Siipien muoto, rakenne ja lukumäärä sekä pigmentin koostumus ja rungon rakenne ovat tärkeitä systemaattisia hahmoja.
Kromuliinijärjestys - Chromulinales. Tähän luokkaan kuuluvat organismit, joilla on monadinen, palmelloidinen ja ameboidityyppinen talluksen erilaistuminen. Monadiset solut, joissa yksi flagellum näkyy valomikroskoopilla.
Suku Chrysameba(Kuva 37, A) - makeanveden ameboidilevät, niiden elinkaaressa on siimavaihe. Tsygoottista kehittyy stomatokysta.
Suku Chromulina(Kuva 37, SISÄÄN) - yksisoluiset vapaasti uivat organismit, jotka elävät pääasiassa makeissa vesissä. Solut ovat alastomia; on yksi tai kaksi kultaista plastidia.
Suku Dinobrion(Kuva 37 , G)- yksisoluiset ja siirtomaa-, vapaasti kelluvat tai kiinnittyneet edustajat. Monadit sijaitsevat maljakkomaisissa taloissa, jotka muodostuvat selluloosamikrofibrillistä. Selluloosan lisäksi talossa on myös suuri määrä aminohappoja. Pesäkkeiden tuuhea muoto liittyy vegetatiivisen solunjakautumisen menetelmään, kun yksi vanhempainkodista lähtevistä tytärsoluista kiinnittyy sen reikään. Pyöriessään akselinsa ympäri se muodostaa oman talonsa. Molemmat lapsisolut voivat myös poistua vanhemmista. Solut sisältävät 1-2 kloroplastia, silmä ja 2 supistuvaa vakuolia näkyvät solun etupäässä. Krysolaminariini sijaitsee tyhjiössä solun takapäässä. Lisääntyminen on vegetatiivista ja seksuaalista.
Tilaa Gibberdiae–Hibberdiales. Tämän luokan edustajille on ominaista ainutlaatuinen pigmenttikoostumus. Fukoksantiinin lisäksi niissä on lisäksi valoa keräävä karotenoidipigmentti, anteraksantiini.
Sisältää synnytyksen Gibberdia, Chromophyton, Styloceras, Chrysopixis, Platyteka ja muut.
Suku gibberdia sen elinkaaressa on kaksi vaihetta: siirtomaa-palmelloidi liikkumaton ja yksisoluinen monadinen liikkuvuus. Valomikroskopialla vain yksi flagellum on näkyvissä monadisessa vaiheessa. Fukoksantiinin lisäksi on ominaista toisen lisävaloa keräävän karotenoidipigmentin, anteraksantiinin, läsnäolo.
Tilaa Hydrorus–Hydrurales. Tämän luokan edustajilla on tallus, jonka rakenne on palmelloidinen ja pseudoparenkymaalinen. Ainutlaatuisten tetraedristen zoosporien esiintyminen on ominaista. Ei ole silmää. Mitoosi on puolisuljettu.
Sisältää synnytyksen Hydrorus, Celloniella, Federmatium, Chrysonebula ja muut.
Suku Hydrurus(Kuva 37, E) on palmelloidilevistä erottuvin. Sen tallus on suuria, jopa 30 cm pitkiä limaisia pesäkkeitä, jotka näyttävät ruskeilta haarautuneilta naruilta, jotka usein lähettävät erittäin epämiellyttävää hajua. Pesäkkeissä voidaan erottaa päärunko ja sivuhaarat. Tavalliseen limaan upotetut solut sisältävät yhden kupinmuotoisen kromatoforin ja useita supistumisvakuoleja. Solut sijaitsevat löyhästi pesäkkeen reunalla ja tiheämpiä keskellä. Tallus voi kasvaa vain jakamalla apikaalisia soluja. Aseksuaalisen lisääntymisen aikana zoosporeja muodostuu pesäkkeen sivuhaarojen soluihin. Muodostaa pallomaisia kystoja. Sitä esiintyy vuoristopuroissa ja joissa, joissa on kylmää vettä, missä se kiinnittyy koviin alustoihin.
Luokka Sinura-levä -Sinurophyceae
syn- yhdessä ja ura- häntä. Tämä organismiryhmä eristettiin vuonna 1987 kultalevistä. Se yhdistää monadisia yksittäisiä ja siirtomaaorganismeja, toisinaan vuorotellen monadisten ja palmelloidisten vaiheiden elinkaaressa. Pektiinikuorten pinta on peitetty piihiutalekuorella. Mitokondriot, joissa on putkimaisia cristae, sijaitsevat yleensä sytoplasmassa kloroplastin ympärillä. Ydin on yksi, kloroplasteja, joita ympäröi neljä kalvoa, yleensä kaksi. Lamellit ovat trityylakoidisia, niissä on vyölamelli. Pääpigmentit ovat klorofyllit A Ja Kanssa, β -karoteeni ja fukoksantiini. Ei ole stigmaa. Solut, joissa on yleensä 2 erilaista siimoa. Pitkä siima on suunnattu eteenpäin. Lyhyt, sileä siima, joskus voimakkaasti kaventunut, suuntautunut taaksepäin.
Solut lisääntyvät pääasiassa pitkittäisjakautumalla. Hajotessaan yhdyskunnat synnyttävät nuoria pesäkkeitä. Joissakin lajeissa seksuaalista prosessia kuvataan isogamiana. Samanaikaisesti, toisin kuin kultalevässä, isogamettien fuusio sinilevissä ei tapahdu etu-, vaan niiden takapäissä. Silicified kystat, joissa on huokoset, muodostuvat endogeenisesti, samanlaisia kuin krysofyyttien.
Pääasiassa planktoniset muodot, palmelloidivaiheet ovat osa pohjaeliöstöä. Useimmat sinuralevälajit ovat fototrofisia makean veden organismeja.
Systematiikka.
Sinura-leväluokkaan kuuluu 7 luokkaa: Chloramoebales, Synurales, Rhizochloridales, Ochromonadales, Heterogloeales, Parmales ja Thallochrysidales.
Tilaa Sinura - Synurales. Monadiset muodot, pektiinisolukalvot on yleensä peitetty piidioksidihiutalekuorella, sementoituna orgaanisella aineella yhdeksi koteloksi. Kloroplastit yksi tai kaksi.
Suku Sinura(Kuva 38) - makeanveden monadiset muodot, joissa on kaksi epätasa-arvoista flagellaa ja kaksi parietaalista kloroplastia. Ydin on päärynän muotoinen, sijaitsee solun etuosassa. Ytimen takana on yksi suuri tyhjiö ja solun takaosassa useita pieniä supistumisvakuoleja. Solut ovat suomujen peitossa, kuten laatat, lisääntyminen on useimmiten vegetatiivista, joidenkin sukupuoliprosessi tunnetaan. Muodostaa pesäkkeitä (kuva 39, A). Usein kystat näkyvät kaikissa pesäkkeen soluissa. Massakehityksellä Sinura voi aiheuttaa veteen epämiellyttävän hajun.

Riisi. 38. Sinura-solujen ulkonäkö
Tilaa Rhizochloride - Rhizochloridales. Pääasiassa makean veden organismeja, joilla on rhizopodiaalinen talluksen erilaistuminen ja jotka elävät pääasiassa makeassa vesistössä.
Suku Mallomonas(Kuva 39, B-D) – yksisoluinen monadi, jossa on yksi hyvin merkitty siima ja yksi kaksihaarainen kloroplasti.

Riisi. 39. Sinuurilevien ulkonäkö (G.A. Belyakova et al., 2006 mukaan): A – halkeavaa siirtokuntaa Sinura; B, C – statosporien muodostuminen ja G – ulkomuoto Mallomonas
Solu on peitetty kaakeloiduilla ja kierteisillä suomuilla, joista osassa on neulamaisia piikkejä (kuva 39, G). Statosporien esiintyminen on ominaista (kuva 39, B, C). Useille edustajille kuvataan seksuaalista prosessia, hologamiaa.
Suku Mixochloris elää sphagnum sammalten lehtien soluissa, näyttää suurelta moniytimiseltä plasmodiumilta. Se muodostaa syksyllä kystat, jotka itävät keväällä. Niistä nousevat eläinporit tai amebat, jotka tunkeutuvat tyhjiin lehtisoluihin ja sulautuvat siellä plasmodiumiksi.
Tilaa Chloramoebaceae - Chloramoebales. Sisältää monadin edustajat. Löytyy suolaisista ja makeista vesistä.
Suku heterochloris – sen solut voivat muuttaa muotoaan muodostaen pseudopodia. Tämä ilmiö on luontainen monille järjestyksen edustajille, samoin kuin taipumukselle eläinravitsemustapaan. Solussa on useita kloroplasteja, öljypisaroita ja krysolaminariinia. Solun edessä on supistuvia vakuoleja, keskellä - yksi ydin. Lisääntyy vegetatiivisesti solujakautumalla.
Tilaa Ochromonadales - Ochromonadales. Yhdistää paljaat muodot kahteen epätasa-arvoiseen siimaan. Makean veden ja meren muodot.
Suku Ochromonas sisältää yksisoluisia monadeja, joissa on kaksi eriarvoista siimat. Solut peitetään vain plasmalemmalla. Solun etupäässä on supistuva vakuoli ja silmä, takapäässä krysolaminariininen tyhjiö. Se lisääntyy vegetatiivisesti, fissioura alkaa solun etupäästä kahden flagellaarisen emäsparin välistä. Suvun lajit ovat yleisiä oligotrofisissa makeissa vesissä, mutta siellä on myös meren edustajia.
Tilaa Heterogleyes - Heterogloeales. Sisältää levät, joilla on palmelloidityyppinen talluksen erilaistuminen. Luokan edustajia tavataan useammin makeissa kuin suolaisissa vesissä.
Suku Helmintoglea asuu murtovesissä. Tämä laajennetulla pohjalla istuva siirtokunta koostuu haarautuvista limaisista nauhoista. Protoplastit ovat satunnaisesti asettuneet tähän limaan, joista jokaista ympäröi oma limakalvo.
Phaeotamnia leväluokka - Phaeothamniophyceae
Luokan nimi tulee tyyppisuvusta Phaeothamnion(kreikasta. phaeos- tumma pensas). Tämän organismiryhmän edustajat tunnistettiin kelta-vihreistä ja kultalevistä erillisessä luokassa vuonna 1998 ribosomaalisten geenien sekvenssin ja solun ultrarakenteen ominaisuuksien analyysin perusteella. Feotamnialevälle on ominaista ainutlaatuinen pigmenttien yhdistelmä: fukoksantiini heteroksantiinin kanssa, violaksantiinin puuttuminen.
Solurakenteen ominaisuudet: vakuolien puuttuminen krysolaminariinilla ja endogeenisten kystien puuttuminen piidioksidiseinämillä.
Luokkaan kuuluu yksisoluisia, siirtomaa- ja monisoluisia organismeja, joilla on kokkoidi-, palmelloidi- ja rihmamainen tallirakenne. Leimautuneet vaiheet, joissa on kaksi eriarvoista siimoa. Flagella lateraalinen tai subapical. Stigmaa löytyy eläinporeista. Pigmentit - klorofyllit A Ja Kanssa, β -karoteeni, fukoksantiini, diadinoksantiini, diatoksantiini ja heteroksantiini. Päävaratuote on paramyloni ( β -glukaani). Soluseinä on läsnä, solunjakautumisen aikana tytärseinä muodostuu emoseinän sisään.
Feotamniaceae-lajeissa tärkeimmistä lisääntymismenetelmistä löytyy vain vegetatiivisia ja aseksuaalisia. Aseksuaalinen lisääntyminen tapahtuu autoitiöiden tai zoosporien avulla. Sukupuolista lisääntymistä ei tunneta.
Feotamnia-levälajit elävät yksinomaan makeissa vesissä. Systematiikka.
Tällä hetkellä feotamnialevien monofyleettinen luokka on vahvistettu, ja okrofyyttien joukossa se on lähempänä ruskea- ja keltaviherleviä kuin kultaleviä. Tällä hetkellä näistä levistä tunnetaan noin 30 lajia, jotka johtuvat yhdestä lahko Theotamnia - Phaeothamniales.
Suku Theotamnion edustaa kiinnitettyjä haarautuvia lankoja, joiden korkeus on enintään 1 cm (kuva 40). Solut ovat sylinterimäisiä, epäsäännöllisiä, ylöspäin laajentuneita

Riisi. 40. Ulkonäkö Theotamnion.
kaksi tai kolme kertaa leveää pidempi. Solunjakautumisen aikana emoseinämän materiaalia kuluu filamenttisoluja ympäröivän kerrostetun vaipan muodostumiseen. Solun reunalla plasmakalvon alla on rakkuloita. Ne muistuttavat fysodeja - muodostumia, joita löytyy ruskealevien soluista. Solut sisältävät yhdestä useisiin kiekon muotoisia oliivinruskeita plastideja. Zoosporeja muodostaa 1-2, harvemmin 4-8 yhdessä solussa. Zoosporeissa flagellat ovat kiinnittyneet sivusuunnassa. Siimajuurten sijainti Theotamnion muistuttaa kelta-vihreitä ja ruskealeviä. Siitä puuttuu myös rhizoplast. Elinkaaren aikana kystat voivat muodostua, mutta niiden seinissä ei ole piitä. Kystat itävät zoosporeina. Laskeutuu rihmamaisten levien epifyyttinä. Asuu pysähtyneissä ja hitaasti virtaavissa makeissa vesistöissä.
Luokka Rafid levät -Raphidophyceae
Luokan nimi tulee kreikan kielestä rafidi - neula. Yhdistää yksisoluisia kaksisoluisia organismeja, joilla ei ole soluseinää. Enimmäkseen makean veden organismeja, joiden solut ovat litistyneet (kuva 41).
Soluilla on eripituisia siimoja, eteenpäin suunnattu siima on pinnallinen ja pidempi, taaksepäin suunnattu siima on sileä ja lyhyempi. Siirtymäkierre puuttuu. Kloroplastit ovat pieniä, lukuisia tai harvemmin 1-2 per solu, ja niitä ympäröi neljä kalvoa, joista kaksi on CES-kalvoja. Tylakoidit on järjestetty kolmen pinoihin. Ydin on suuri, ja sitä ympäröi diktyosomirengas. Mitokondriot putkimaisilla risteillä. Siellä on supistuvia vakuoleja. Sytoplasma on usein vakuoloitunut. Solut ovat alastomia, ja niitä ympäröi vain plasmalemma. Stigma (silmä) puuttuu yleensä.
Fotosynteettiset pigmentit - klorofyllit A Ja Kanssa, β -karoteeni, vosheriaksantiini, dinoksantiini, diadinoksantiini, heteroksantiini.
Vara-assimilaatiotuotteita ovat rasvat ja öljyt, harvemmin tärkkelys ja krysolaminariini.
Ne elävät myös sfagnum-soissa ja muissa elinympäristöissä, joissa on happamia ja neutraaleja vesireaktioita. Jotkut lajit löytyvät murto- ja merivesistä.

Riisi. 41. Rapidlevien ulkonäkö.
Systematiikka.
Hyvin pitkään tätä leväryhmää pidettiin sen vihreän värin, nielun läsnäolon ja useiden muiden rakenteellisten ominaisuuksien vuoksi euglenoidien joukossa Chloromonas-luokkana. Mutta sytologian, biokemian, fysiologian ja molekyylibiologian tiedot osoittivat niiden kiistattoman kuulumisen Heterocont (Ochrophyte) leväjakoon ja Rafid-leväryhmän monofyleettisen luonteen.
Ainoastaan tunnetaan noin 25 lajia tilaa Huttonellaceae– Chattonellales.
Suku heterosigma(Kuva 42, A) sisältää merisiimat. Soikeat solut, joissa flagella on siirtynyt sivulle, plastidit sijaitsevat solun reunalla. Trikokystiä ei ole. Ne uivat ja tekevät pyöriviä liikkeitä kehon pituusakselin ympäri. Saattaa aiheuttaa myrkyllisiä "vesikukintoja" rannikkovesissä.

Riisi. 42. Rafid levät (tekijä: R. E. Lee, 1999): A – heterosigma; B – Goniostomum; 1 - kloroplasti; 2 - mukokysti; 3 - supistuva vakuoli; 4 - trikokysti; 5 - ydin
Suku Goniostomum(Kuva 42, B) ovat liikkuvia dorsoventraalisesti puristettuja monadeja. Selkäpuoli on kaareva, vatsapuoli litistynyt. Etupäässä oleva uurre johtaa kolmiomaiseen nieluun, josta kaksi pitkää siimat nousevat esiin; niiden pituus on verrattavissa solun pituuteen. Plastidit sijaitsevat solun reunalla. Plasmalemman alla on trikokystoja. Asuu makeissa vesissä, joiden pH on hapan.
Luokka Eustigma levät -Eustigmatophyceae
Luokan nimi tulee kreikan kielestä eu- "hyvä ja stigma- "merkki", "piste". Luokka yhdistää paljaat yksisoluiset, harvemmin siirtomaa-organismit, joilla on pääosin kokkoidirakenne.
Yksi tai useampi ydin. Kloroplasti on yleensä yksi, jota ympäröi 4 kalvoa. Siima on yleensä yksi etuosa ja toinen tyvirunko tai harvemmin kaksi epätasaista siipiä. Stigma (silmä) on läsnä, sijaitsee kloroplastin ulkopuolella, sen rakeet ovat ilman kalvoja.

Riisi. 43. Eustigma-levien ulkonäkö
Kloroplasti on yleensä yksittäinen, suuri, moniliuskainen, kuppimainen tai parietaalinen, väriltään kelta-vihreä. Tylakoidit on ryhmitelty lamelleiksi kolmen ryhmän ryhmiin; ympäröivä lamelli puuttuu.
Pääpigmentit ovat klorofylli a, β-karoteeni, violaksantiini ja vosheriaksantiini. Pyrenoidi esiintyy yleensä vain vegetatiivisissa muodoissa.
Vara-aineet ovat öljyjä ja luonteeltaan tuntemattomia aineita, jotka kerrostuvat kiinteänä aineena plastidien ulkopuolelle. Tärkkelystä ei muodostu.
Ne elävät makeissa vesissä, ovat vähemmän yleisiä merissä ja maaperässä.
Systematiikka.
Aikaisemmin tämä organismiryhmä luokiteltiin keltavihreiksi leviksi. Tällä hetkellä Eustigma-leviä pidetään luokkana Heterocontophytes-osastolla. Noin 35 lajia tunnetaan, enimmäkseen makean veden, katsotaan ainoaksi Eustigma järjestys - Eustigmatales.
Suku Eustigmatos(Kuva 43) - yksisoluiset pienet pallomaiset levät. Soluseinä on sileä, ehjä, ilman koristeita. Keltaisenvihreä kloroplasti yksittäinen, lohko, parietaalinen, varrellisia monitahoisia pyrenoideja. Suuri keskusvakuoli punaisella sisällöllä. Se lisääntyy kahdella tai neljällä autoitiöllä tai pullon muotoisella zoosporilla, joissa on yksi sisääntuleva siima, suuri stigma, joka sijaitsee kloroplastin ulkopuolella, etupäässä ja yksi posteriorinen kloroplasti ilman pyrenoidia. Maaperän kasviston yleinen komponentti. Tunnetaan Uudesta-Seelannista, Itävallasta, Islannista ja Arizonan altailta "sinappimerilevänä".
Luokka Kelta-vihreät levät -Xanthophyceae
Keltavihreät levät ovat niitä, joiden kloroplastit ovat vaalean tai tummankeltaisia, hyvin harvoin vihreitä ja vain joskus sinisiä. Tallin värin määrää seuraavien pigmenttien läsnäolo kloroplastisoluissa - klorofyllit A Ja Kanssa, β -karoteeni ja ksantofyllit. Jälkimmäisen hallitsevuus määrittää kelta-vihreiden levien värin omaperäisyyden. Lisäksi paramylonia, öljypisaroita ja vain joissakin lajeissa lisäksi leukosiinin ja volutiinin kokkareita kerääntyy soluihin assimilaation päätuotteena. Keltavihreät levät eivät muodosta tärkkelystä. Keltaisen vihreän erottuva piirre on monadisen rakenteen läsnäolo vegetatiivisissa soluissa ja kaksi epätasa-arvoista flagellaa zoosporeissa. Soluseinä sisältää selluloosaa, glukoosia ja uronihappoja. Soluseinä koostuu usein kahdesta osasta.
Lisääntyminen on vegetatiivista, aseksuaalista ja seksuaalista.
Levitetty laajalti makeissa vesissä. Harvoin meri-, murtovesi- ja maaperässä.
Aikaisemmin keltavihreäleväluokkaa kutsuttiin Tribophytium leväksi Tribonema-tyyppisuvun mukaan (kreikasta. tribon – taitava, ovela ja nema – lanka). Noin 450 lajia tunnetaan.
Keltaisille vihreille on ominaista merkittävä morfologinen monimuotoisuus. Tämän osaston lukuisten edustajien joukosta löytyy melkein kaikki päätyypit kehon rakenteesta: ameboidi, monadinen, palmelloidi, kokkoidi, rihmamainen, monisäikeinen, lamellaarinen ja sifonaalinen (kuva 44). – 46). Tallus yksisoluinen,

Riisi. 44. Kelta-vihreiden levien ulkonäkö: 1, 2 - Charatiopsis, 3 – Centritractus, 4 – ofiosytium
siirtomaa, monisoluinen ja ei-soluinen. Solukalvo on tiivistä, pektiinistä ja selluloosasta, joka koostuu tiiviisti limittäisistä osista tai kahdesta venttiilistä. Piidioksidia tai kalkkia kertyy kuoreen. Enimmäkseen kiinteät muodot. Yksisoluisten lajien joukossa on liikkuvia muotoja, joissa ei ole tiheää kuorta ja jotka on varustettu flagellalla, lobopodialla ja rhizopodialla.

Riisi. 45. Ksantokokin keltavihreän levien ulkonäkö: 1-3 - Botrydiopsis, 4 – tetrahedriella, 5 – Pseudostaurastrum, 6 – Goniochloris, 7, 8 – Bumilleriopsis
Useimmat kelta-vihreät – liikkumattomat organismit. Liikkuvilla yksilöillä liikkeet voidaan suorittaa käyttämällä flagellaa tai rhizopodiaa. Erimuotoisia soluja: pallomaisia, karan muotoisia, ellipsoidisia, sylinterimäisiä, tetraedrisä, sirpin muotoisia, päärynän muotoisia, munamaisia. Talluksen koot alkaen 0,5 – 1,5 µm ( Chloridella) halkaisijaltaan useita millimetrejä ( Botrydiopsis) (Kuva 45, 1 – 3) ja jopa kymmeniä senttejä pitkä ( Vosheria) (Kuva 46, 3).

Riisi. 46. Kelta-vihreiden levien ulkonäkö: 1 - Tribonema, 2 – heteropedia, 3 – Vosheria, osa filamenttia, jossa on oogoniumia ja anteridiumia
Suurin osa keltavihreistä lajeista on fototrofeja, mutta myös holotsoista ravintoa esiintyy bakteerien ja pienten levien nauttimisen kautta. Keltalevät ovat laajalle levinneitä makeissa vesissä. Ne ovat yleisiä myös maaperässä, vähemmän yleisiä meri- ja murtovesissä. Luokka sisältää aerobiontti-, plankton-, pohja- ja perifytoniset muodot. Epifyytit, epitsoiitit sekä solunsisäiset symbiontit alkueläinsoluissa.
Ulkoisesta rakenteesta riippumatta kelta-vihreäleväsolun sisäinen rakenne on sama. Protoplastissa on yleensä useita kellanvihreitä kloroplasteja, jotka ovat levymäisiä, kaukalomaisia, lamellisia, harvemmin nauhamaisia, tähtimäisiä tai kuppimaisia kokonaisia tai lohkoreunaisia. Väritys johtuu fukoksantiinin puuttumisesta, joka on vastuussa muiden okrofyyttien kultaisesta ja ruskeasta väristä. Niistä muista pigmenteistä β -karoteeni, vosheriaksantiini, diatoksantiini, diadinoksantiini, heteroksantiini. Liikkuvissa muodoissa punasilmäisyys tai stigma sijaitsee yleensä kloroplastin etupäässä. Harvalla lajilla on puoliksi upotettuja pyrenoideja. Solun ydin on yleensä yksi, pienikokoinen, mutta on lajeja, joissa on monitumaisia soluja. Joillakin lajeilla on yksi tai kaksi supistuvaa (sykkivää) vakuolia solun etuosassa.
Monadien edustajilla ja liikkuvilla vaiheilla (eläinporit ja sukusolut) on kaksi epätasa-arvoista siimat. Poikkeuksena ovat synzoosporit. Vosheri, jossa pinnalla on lukuisia sileitä, pituudeltaan hieman erilaisia siimoja. Lyhyt flagellum päättyy akroneemiin. Siimat kiinnittyvät soluun subaapisesti. Siittiössä Vosheri sivukiinnitys.
Lajeissa, joissa on ameboidi-, monadaalinen- ja palmelloidiorganisaatio, soluseinä puuttuu, niitä peittää vain sytoplasminen kalvo ja ne voivat helposti muuttaa muotoaan. Joskus "paljaita" soluja löytyy taloista, joiden seinät voidaan maalata ruskeiksi mangaani- ja rautasuoloilla. Suurimmalla osalla muotoja on soluseinä, joka koostuu kahdesta osasta. Selluloosa vallitsee soluseinän koostumuksessa, ja se sisältää myös polysakkarideja, jotka koostuvat pääasiassa glukoosista ja uronihapoista. Nuorissa soluissa kalvo on ohut, iän myötä se paksunee. Siihen voi kertyä rautasuoloja, joiden yhdisteet värjäävät sen ruskean ja punaisen eri sävyissä. Piidioksidia on useammin soluseinässä, mikä antaa sille kovuutta ja kiiltoa. Se voidaan myös peittää kalkilla ja muotoilla eri tavoin (piikit, solut, syyliä, syyliä, hampaat jne.) Kuoren uloskasvu voi muodostua kiinnittyneissä muodoissa – jalka kiinnityspohjalla.
Kelta-vihreälevien rihmamaisissa muodoissa, joissa on simpukoita, solukalvot hajoavat filamenttien hajottua H-muotoisiksi fragmenteiksi. Nämä fragmentit ovat tiiviisti yhdistettyjä kahden vierekkäisen solun kuoren puolikkaita (kuva 47). Filamenttien kasvun aikana kahden vierekkäisen tytärsolun soluseinän H-muotoinen fragmentti on upotettu emosolun kalvon kahden puolikkaan väliin. Tämän seurauksena jokainen tytärsolu on puoliksi peitetty emosolun vanhalla kalvolla ja puoliksi – vasta muodostettu kuori.

Riisi. 47. Kaavio poikittaisen väliseinän muodostumisesta kahden tytärsolun välille rihmamaisissa keltavihreissä levissä (A.A. Masyukin, 1993 mukaan): A- langan fragmentti; B- vyörenkaan asettaminen ja poikittaisen väliseinän muodostaminen kahden solun välille; SISÄÄN– simpukoiden solukalvojen kerrostaminen; G– kuoren hajoaminen H:n muotoisiin osiin
Liikkuvissa edustajissa on supistuvat tyhjiöt. Yleensä niitä on 1-2 per solu. Omalaatuisen rakenteen Golgi-laite. Diktyosomit ovat pieniä, sisältävät 3-7 säiliötä. Ydin on yksi, harvemmin niitä on useita; koenoottisissa lajeissa solut ovat aina moniytimiä.
jäljentäminen. Useimmille keltavihreille levälajeille on ominaista kasvullinen ja suvuton lisääntyminen.
Vegetatiivinen lisääntyminen toteutetaan eri tavoin: solujen jakautuminen puoliksi, pesäkkeiden romahtaminen ja monisoluinen talli osiin. klo Vosheri muodostuu erityisiä pesäsilmuja.
klo suvuton lisääntyminen Erilaisia itiöitä voi muodostua: amoeboideja, zoosporeja, synzoosporeja, autoitiöitä, hemisoosporeja, hemiautosporeja, aplanosporeja. Zoosporit ovat "alastomia" ja yleensä päärynän muotoisia.
seksuaalinen prosessi- isogamia, heterogamia ja oogamia - kuvattu muutamassa edustajassa. klo Tribonemes Sukusolut ovat kooltaan samanlaisia, mutta käyttäytyvät erilaiset - tämä on isogamiaa. klo Vosheri Oogamiaa havaitaan: lankoihin muodostuu naarassukusolujen astioita – oogonia ja uros – anteridia.
Epäsuotuisissa olosuhteissa havaitaan kystan muodostumista. Kystat (statosporit) ovat endogeenisiä, yksitumaisia, harvoin moniytimiä. Niiden seinämä sisältää usein piidioksidia ja koostuu kahdesta epätasaisesta tai harvemmin yhtä suuresta osasta.
Systematiikka.
XIX vuoden lopussa – 1900-luvun alku Viherleväosastolle sijoitettiin useita keltavihreäleväsuvuja, jotka liittyivät ensisijaisesti tallin väriin ja morfologiseen samankaltaisuuteen. Keltaisia vihreitä pidetään tällä hetkellä okrofyyttiluokkaan kuuluvana luokkana.
Keltavihreäleväluokan nykyaikaisia lajeja tunnetaan noin 450, jotka on ryhmitelty neljään luokkaan: Botridia, Mishococcus, Tribonem ja Vosheria. Tilausten luokittelu perustuu talluksen erilaistumistyyppiin ja elinkaaren ominaisuuksiin.
Tilaa Botridiae - Botrydiales. Luokka sisältää lajeja, joilla on sifonaalinen talluksen erilaistuminen, jossa ei ole oogaamia sukupuoliprosessia.
Suku Botridium elää maaperässä ja näyttää muutaman millimetrin kokoisilta vihreiltä kuplilta, joihin on kiinnitetty värittömiä risoideja. Tallus on sifonaalinen, sisältää lukuisia ytimiä ja plastideja. Kuori on monikerroksinen, sen päälle voidaan kerrostaa kalkkia. Lisääntyminen on suvutonta biflagellate zoosporien avulla, kun taas rakon koko sisältö hajoaa yksitumaisiksi fragmenteiksi. Kosteuden puutteessa se lisääntyy aplanosporien avulla tai muodostaa paksuseinäisiä kystoja. Joissakin tapauksissa koko virtsarakon sisältö menee yhden suuren kystan muodostumiseen. Muissa tapauksissa kystat muodostuvat risoideihin, joissa virtsarakon sisältö ensin kulkeutuu. Kystat itävät joko suoraan uuteen tallukseen tai muodostavat zoosporeja. Seksuaalinen prosessi on iso- ja heterogamia. Tsygootti itää välittömästi, ilman lepojaksoa. Yleisiä ja laajalle levinneitä lajeja maa-elinympäristöissä, joita tavataan purojen, lampien rannoilla tai maaperällä, jossa ei ole kasvillisuutta.
Tilaa Mischococcal–Mishococcaleja. Yksisoluiset tai koloniaaliset edustajat, joilla on kokkoidityyppinen talluksen erilaistuminen.
Suku Charatiopsis sisältää yksisoluiset kiinnittyneet muodot. Lisääntyessään se muodostaa zoosporeja, aplanosporeja ja paksuseinäisiä kystoja (Kuva 44, 1-2).
Suku ofiosytium(Kuva 44, 4) on pitkänomaiset lieriömäiset kennot, jotka voivat olla suoria, taivutettuja tai spiraalimaisesti kierrettyjä ja joissa voi olla piikki. Soluseinä koostuu kahdesta epätasaisesta osasta, joista suurin osa osallistuu solujen kasvuun, pienempi osa on pysyvää ja korkin muotoista. Yksisoluinen ja siirtomaalaji, vapaasti elävä tai kiinnittynyt alustaan pienellä varrella. Ne lisääntyvät zoosporien ja aplanosporien avulla, kystat löytyvät. He elävät makeissa vesissä.
Suku Mischococcus muodostaa puumaisia kiinnittyneitä pesäkkeitä. Haaroittuminen on kaksijakoista ja tetrakotomista. Solut sijaitsevat 2 tai 4 pesäkkeen limahaarojen yläosissa. Solut ovat pallomaisia tai soikeita, ja niissä on ohut tai paksu soluseinä. Joskus soluseinä on kiiltävä ja ruskea, koska se on kyllästetty rautasuoloilla. Nuoret yksisoluiset organismit, joilla on limainen, kiekon muotoinen pohja, joka toimii kiinnittymisen tukipisteenä. Itiöiden vapautumisen jälkeen emosolun protoplasti muuttuu hyytelöksi ja venyy, pituus on 6 kertaa suurempi kuin leveys, ja siten syntyy lieriömäinen jalka. Emosolun tyhjä soluseinä tulee aina kantasolun pohjaksi. Aseksuaalinen lisääntyminen zoosporien ja autosporien avulla. Autoitiöt ovat kiinnittyneet limakalvon varren yläreunaan. Seuraavat solujakautumat toistavat prosessin ja tuottavat puun kaltaisen pesäkkeen. seksuaalinen prosessi – isogamia. Ne elävät pienissä makeissa vesistöissä rihmamaisten levien epifyytteinä. Tunnettu Keski-Euroopassa ja Aasiassa.
Tilaa Tribonemaceae - Tribonematales. Edustajilla on rihmamainen, monisäikeinen, väärän kudoksen ja kudostyyppinen talluksen erilaistuminen. Soluseinät ovat joko H-muotoisia päällekkäisiä osia tai kiinteät.
Suku Tribonema- haarautumattomat kierteet (kuva 46, 1). Solut ovat sylinterin tai tynnyrin muotoisia. Soluseinä koostuu kahdesta puolikkaasta, joiden reunat ovat päällekkäin solun keskellä. Kuoret ovat usein kerroksittain. Lankojen palaset päättyvät aina tyhjiin H-muotoisten kuorikappaleiden puolikkaisiin, jotka näyttävät haarukalta. Soluissa on useita kellertävänvihreitä plastideja, pyrenoideja ei ole. Lisääntyminen on vegetatiivista (filamenttien sirpaloituminen), aseksuaalista (zoosporien ja aplanosporien kautta) ja sukupuolista (isogamia), ja aplanosporeja tuotetaan useammin kuin zoosporeja. Voi muodostaa akinetteja. Ne elävät makeissa vesissä, joissa ne kehittyvät erityisen runsaasti kylmänä vuodenaikana.
Tilaa Vosherievye - Vaucheriales. Kaikilla edustajilla on sifonaalinen tallus, oogaminen seksuaalinen prosessi ja synzoosporit.
Suku Vosheria(kuvio 46, 3) on tallus, jolla on ei-solurakenne; sen tallus saavuttaa useiden senttimetrien pituuden, kiinnittyy alustaan värittömän risoidin avulla. Langoissa ei ole väliseiniä, suurin osa talluksesta on tyhjiössä, ja lukuisat ytimet ja plastidet sijaitsevat sytoplasman reunalla. Langat, joissa on apikaalinen kasvu ja harvinainen sivuhaaroitus. Väliseinät muodostuvat, kun talli on vaurioitunut ja erottamaan sukuelimet. Aseksuaalista lisääntymistä suorittavat aplanosporit, synzoosporit, akinetit. Synzoosporit muodostuvat yksi kerrallaan zoosporangiumissa, joka on erotettu vegetatiivisista soluista filamentin päässä olevalla väliseinällä. Eläinporit ovat monitumaisia ja monisiimaisia. Seksuaalinen prosessi on oogamiaa. Tsygootti on peitetty paksulla kuorella ja kasvaa lepotilan jälkeen uudeksi tallukseksi.
Erilaisia Vosheri levinnyt laajalti makeissa, murto- ja merivesissä sekä maalla olevissa elinympäristöissä. Niitä löytyy kaikilla mantereilla, myös Etelämantereella. Ne muodostavat ruohonvihreitä tai tummanvihreitä toisiinsa kietoutuneita massoja - niin sanottuja mattoja, sileitä, hiipiviä tai tyynynmuotoisia. Vesi-, puolivesi-, maanpäälliset muodot. Ne elävät erilaisissa elinympäristöissä: merissä, suistoissa, suistoissa, suolamaissa, mangrovemetsissä, puroissa, kanavissa, järvissä, lammissa, peltomailla ja soilla.
Merkitysheterokont levät
Heterocont-leväosastolta ruskealevällä on suurin merkitys luonnollisille ekosysteemeille ja ihmisille.
Ruskeat levät - tärkein orgaanisen aineen lähde merten rannikkovyöhykkeellä. Niiden biomassa lauhkean ja subpolaarisen vyöhykkeen merissä voi olla useita kymmeniä kiloja neliömetriä kohti. Ruskealeväpaksut tarjoavat olosuhteet monien rannikkoeläinten ja muiden levien ruokimiselle ja lisääntymiselle. Charles Darwin tarkkailee ruskealevää Etelä-Amerikan rannikolla Makrocystis, kirjoitti: "Voin vain verrata näitä eteläisen pallonpuoliskon valtavia vedenalaisia metsiä trooppisten alueiden maanpäällisiin metsiin. Ja silti, jos jossain maassa tuhoutuisi metsä, en usko, että edes suunnilleen sama määrä eläinlajeja kuolisi kuin tämän levän tuhoutuessa.
Ruskealevän paksut palvelevat ruokapaikka,suojaa ja kasvatus monia eläimiä. Kuvaannollisesti sanottuna ruskeat levät tarjoavat muille vesieliöille "pöydän, suojan ja lastenhuoneen".
Ruskealevää käytetään laajalti ihmisillä. He ovat rikkaita jodi ja muita mikroravinteita. Kaakkois-Aasian kansat käyttävät niitä perinteisesti ruoaksi, erityisesti rakkolevälajin edustajia, joista valmistetaan monenlaisia ruokia. Syö ateria ruskealevistä valmistettu, lisää karjan tuottavuutta; Samalla munien ja maidon jodipitoisuus kasvaa.
Ruskeista levistä vastaanottaaalginaatit- algiinihapon suolat. Alginaatteja käytetään laajasti eri teollisuudenaloilla. Nämä ovat myrkyttömiä yhdisteitä, joilla on kolloidisia ominaisuuksia, joten niitä käytetään laajalti elintarvike- ja lääketeollisuudessa. Algiinihappo ja sen suolat kykenevät 200-300-kertaisesti imemään vettä muodostaen geelejä, joille on ominaista korkea haponkestävyys. Elintarviketeollisuudessa niitä käytetään pääasiassa emulgointiaineina, stabilointiaineina, hyytelöimis- ja vettä pidättävinä komponentteina. Esimerkiksi kuivajauhenatriumalginaattia käytetään jauhettujen ja briketoitujen liukenevien tuotteiden (kahvi, tee, maitojauhe, hyytelö jne.) valmistuksessa niiden nopeaan liukenemiseen. Alginaattien vesiliuoksia käytetään liha- ja kalatuotteiden pakastamiseen. Jopa 30 % maailmassa tuotettujen alginaattien kokonaismäärästä menee elintarviketeollisuuteen.
Tekstiili- ja massa- ja paperiteollisuudessa alginaatteja käytetään maalin sakeuttamiseen ja niiden sidoksen lujuuden lisäämiseen pohjan kanssa. Kankaiden kyllästäminen joillakin alginaateilla antaa niille suojaavia ominaisuuksia: vedenpitävyyttä, haponkestävyyttä ja lisää mekaanista lujuutta. Keinotekoisen silkin valmistamiseen käytetään lukuisia algiinihappojen suoloja. Toisen maailmansodan aikana Yhdysvalloissa ja Englannissa valmistettiin algiinihaposta ja sen suoloista suuri määrä naamiointikankaita ja verkkoja asuin- ja teollisuusrakennuksiin.
Alginaatteja käytetään metallurgiassa: valimossa ne parantavat valumaan laatua. Algiinihapon suoloja käytetään sähköhitsauksen elektrodien valmistuksessa, mikä mahdollistaa laadukkaampien hitsausten valmistamisen. Alginaatteja käytetään myös muovien, synteettisten kuitujen, pinnoitteiden ja säänkestävien rakennusmateriaalien valmistuksessa. Niitä käytetään korkealaatuisten koneiden voiteluaineiden valmistukseen. Radioelektroniikassa alginaatit toimivat sideaineena korkealaatuisten ferriittien valmistuksessa.
Suurin sovellus löytyy vesiliukoisesta natriumalginaatista, joka pystyy muodostamaan viskoosisia liuoksia. Sitä käytetään laajasti erilaisten liuosten ja suspensioiden stabilointiin. Pienen määrän natriumalginaattia lisäämällä elintarvikkeisiin - säilykkeisiin, jäätelöön - niiden laatu paranee. Siitä valmistetaan myös koristekosmetiikkaa, voiteita ja naamioita hajuvesiteollisuudessa.
Lääketeollisuudessa alginaatteja käytetään tablettien, pillereiden päällystämiseen, erilaisten voiteiden ja tahnojen komponenttipohjana, lääkekantajageeleinä sekä liukenevien kirurgisten ompeleiden valmistuksessa. Lääketieteessä kalsiumalginaattia käytetään hemostaattisena aineena ja radionuklideja (esimerkiksi strontiumia) poistavana sorbenttina. Alginaattien vuosituotanto maailmassa ylittää 20 tuhatta tonnia.
Toinen tärkeä ruskealevistä peräisin oleva aine on kuusiarvoinen alkoholimannitoli. Mannitolia käytetään sokerinkorvikkeena diabeetikoille. Lisäksi sitä voidaan käyttää plasman korvikkeena veren säilönnässä. Sitä käytetään tablettien valmistukseen lääketeollisuudessa. Mannitolia käytetään myös mm synteettisten hartsien, maalien, paperin, räjähteiden, nahkasidosten tuotanto.
fukoidaanit, ruskealevistä saadut ovat tehokkaita antikoagulantteja, jopa hepariinia aktiivisempia. Niiden käyttöä kasvainlääkkeiden ja viruslääkkeiden valmistukseen pidetään lupaavana. Loppujen lopuksi, jopa pienimmällä pitoisuudella, fukoidaanit voivat estää virusten kiinnittymisen solujen pintaan. Fukoidaaneilla on myös kyky muodostaa poikkeuksellisen vahvaa ja viskoosia limaa, jota käytetään stabiilien emulsioiden ja suspensioiden valmistukseen.
Monia maailman maita viime vuosina valtannut energiakriisi on johtanut tarpeeseen etsiä uusia ei-perinteisiä energialähteitä. Joten Yhdysvalloissa tätä tarkoitusta varten tutkitaan mahdollisuutta kasvattaa jättiläisleväleviä. Makrocystis jonka jälkeen se prosessoidaan metaaniksi. On laskettu, että tämän levän 400 km 2:n alueelta voidaan saada 620 miljoonaa m 3 metaania.
Heterokonttilevät luokista Golden, Yellow-green, Sinura, Rafidophyte ja Eustigma, joita edustavat pääasiassa mikroskooppiset organismit, ovat yleisiä makeissa vesistöissä kaikilla maapallon ilmastovyöhykkeillä, mutta ovat yleisempiä lauhkeilla leveysasteilla. Kultalevien joukossa on lajeja, jotka elävät merissä ja suolajärvissä, ja hyvin harvat saastuneissa vesissä. Kultalevät saavuttavat huippukehityksensä kylmänä vuodenaikana: ne hallitsevat planktonissa aikaisin keväällä, myöhään syksyllä ja talvella. Tällä hetkellä niillä on merkittävä rooli alkutuotannon tuottajina ja ne toimivat eläinplankton-organismien ravinnoksi.
Jotkut kultalevät esimerkiksi Uroglena Ja Dinobrion, kehittyvät massamäärinä, ne voivat aiheuttaa vesikukintoja. Ne vapauttavat aldehydejä ja ketoneja, jotka voivat antaa vedelle epämiellyttävän hajun ja maun, a Uroglena– kalalle myrkyllisiä rasvahappoja.
Rafid-levät ovat laajalti edustettuina pääosin happaman pH:n makean vesistön planktonissa, erityisesti sfagnum-soissa, harvemmin suurissa järvissä. Makeissa vesistöissä voi muodostua paikallisia "kukintoja". Goniostomum. Rafid-leviä esiintyy myös suolattomissa merenlahdeissa ja lätäköissä merenrannalla sekä avomerellä. Rannikon merivesien massojen kehittyessä ne aiheuttavat myrkyllistä veden "kukintoa". Joten Kanadan rannikolla "kukinnan" aikana Heterosigma-rafid-levän solujen pitoisuus voi nousta 30 miljoonaan litrassa. Rapidlevien puhkeaminen johtaa usein "punaisten vuorovesien" kehittymiseen, jotka liittyvät kalojen kuolemaan. Tällaisten "punaveden" syy voi olla synnytyslajit Hattonella, Olistodiscus, Heterosigma ja Fibrocapsa.
Makeissa vesistöissä massoittain kehittyneet sinuralevät voivat aiheuttaa veteen epämiellyttävän hajun ( Sinura). Feotamnialeviä esiintyy pysähtyneissä ja hitaasti virtaavissa makeissa vesistöissä, joissa ne asettuvat epifyyttisesti rihmaleville.
Eustigma-levää esiintyy vain makeassa vedessä tai maaperässä.
Keltaleviä on levinnyt kaikille mantereille, ne elävät pääasiassa makeassa vedessä ja maaperässä sekä maa-, murtovesi- ja meriympäristöissä. Keltalevät elävät puhtaissa ja saastuneissa vesissä, joiden pH-arvot vaihtelevat: ne voivat elää sekä happamissa että emäksissä. Niitä esiintyy pääasiassa puhtaissa makean veden säiliöissä, harvemmin merissä ja murtovesissä, mieluummin maltillisia lämpötiloja, kehittyvät useammin keväällä ja syksyllä, vaikka on lajeja, joita esiintyy kaikkina vuodenaikoina, myös talvella. Useimmiten niitä löytyy lankaryhmistä ja korkeampien vesikasvien pensaikkoista jokien, lampien, järvien ja tekoaltaiden rannikkoalueilla.
Suurin osa keltavihreistä on vapaasti eläviä muotoja, mutta alkueläinsoluissa on myös solunsisäisiä symbiontteja - zooksanteeleja. Meren lajien kloroplastit muodostavat mielenkiintoisen solunsisäisen symbioosin Vosheri simpukan kanssa Elisio. Yhdeksän kuukauden ajan tämä nilviäinen pystyy kiinnittämään fotoautotrofisesti hiilidioksidia viljelmässä. Tämä on pisin tämän tyyppinen symbioosi, kun symbioottinen plastidi on suorassa kosketuksessa eläimen sytoplasmaan. Luonnossa nilviäisten toukat ruokkivat lankoja. Vosheri. Fagosytoosin seurauksena leväkloroplastit pääsevät nilviäisten epiteelisolujen sytoplasmaan. Tämän prosessin aikana kloroplastikalvo muuttuu kolmikerroksiseksi, ja yksi CES:n (kloroplastin endoplasminen retikulum) ulkokalvo menetetään. Tämä ilmiö on hyvä todiste siitä, että evoluution aikana kolme kalvoa sisältäviä kloroplasteja voi syntyä sekundaarisen symbiogeneesin seurauksena kalvojen häviämisen seurauksena.
Keltavihreät, kultaiset ja muut heterokonttilevät ovat hapen ja orgaanisten aineiden tuottajia; ne ovat osa ravintoketjua. Heterokontofyytit osallistuvat saastuneiden vesien ja maaperän itsepuhdistumiseen, sapropeelin muodostumiseen, orgaanisen aineksen kertymiseen maaperään, mikä vaikuttaa sen hedelmällisyyteen. Niitä käytetään edustavina organismeina määritettäessä veden saastumisen tilaa; keltavihreät levät ovat osa jäteveden käsittelyyn käytettyä mikro-organismikompleksia.
Kontrollikysymykset
Nimeä ruskealevän rakenteen ominaispiirteet.
Ruskealeväthallin rakenteelliset piirteet.
Kuinka ruskeat levät lisääntyvät? Mitä ovat monosporit, tetrasporit ja zoospores, isogamia, heterogamia ja oogamia?
Mitkä ovat ruskealevien elinkaaret? Fucus- ja rakkolevän lisääntyminen.
Nimeä ruskealevälajien tunnusmerkit ja tyypilliset edustajat.
Missä elinympäristöissä ruskealeviä löytyy? Mikä on niiden merkitys luonnossa.
Ruskealevien taloudellinen merkitys.
Nimeä kultalevän rakenteen ominaispiirteet ja tyypilliset edustajat.
Mitä pigmenttejä ja ravintotyyppejä kultalevistä tunnetaan?
Kultalevien lisääntyminen ja ekologia.
Nimeä kelta-vihreiden levien tunnusmerkit ja tyypilliset edustajat.
Mitä pigmenttejä ja ravintotyyppejä keltalevissä tunnetaan?
Miten keltavihreät lisääntyvät? Sukupuolisen lisääntymisen tyypit: isogamia, heterogamia ja oogamia?
Nimeä sinilevän rakenteen ominaispiirteet ja tyypilliset edustajat.
Sinuurilevien lisääntyminen ja ekologia.
Nimeä feotamnialevien rakenteen ominaispiirteet ja tyypilliset edustajat.
Feotamnialevien lisääntyminen ja ekologia.
Nimeä rafidofyyttilevien rakenteen yleiset piirteet ja tyypilliset edustajat.
Rafidophyte levien lisääntyminen ja ekologia.
Mitkä ovat Eustigma-levien rakenteen ja tyypillisten edustajien yhteiset piirteet.
Eustigma-levien lisääntyminen ja ekologia.
Heterokontlevien arvo luonnollisissa ekosysteemeissä.
Flagella - vanhin alkueläinryhmä, joka sijaitsee eläinmaailman sukupuun juuressa ja yhdistää eläimet kasvimaailmaan. Vielä vuosisadan puolivälissä kuuluisa venäläinen biologi L. S. Tsenkovsky (1822-1887) huomautti terävän rajan puuttumisesta siimautuneiden alkueläinten ja yksisoluisten levien välillä. Todellakin, jotkut siimalevät (euglenoidit ja fytomonadit) luokitellaan perustellusti sekä alkueläintyypeiksi, eli eläimiksi, että viherlevätyypeiksi, eli kasveiksi.
Luokan ominaisuus
Yksi tärkeimmistä alkueläinten luokittelussa käytetyistä ominaisuuksista ovat liikeorganellit. Siimalilla nämä ovat nippuja tai vitsauksia - ohuita sytoplasmisia kasvaimia. Ne sijaitsevat yleensä kehon etupäässä ja syntyvät erityisistä tyvijyväistä (tyvikappale tai kinetosomi). Siimaleiden lukumäärä eri lajeissa vaihtelee 1-8 tai enemmän. Joissakin siimoissa (leishmania, trypanosomes) siiman tyveen sijoitetaan lisäksi erityinen organoidi, kinetoplast. Ultrarakenteeltaan se vastaa mitokondrioita, mutta erottuu korkeasta DNA-pitoisuudesta. Uskotaan, että kinetoplastissa syntyy energiaa siiman liikettä varten, joka suorittaa pyörivän liikkeen ja ikään kuin ruuvataan veteen. Joillakin luokan edustajilla flagellum kulkee kehoa pitkin yhdistäen siihen ohuella sytoplasman kasvulla. Mainittu uloskasvu eli aaltoileva kalvo suorittaa aaltoilevia liikkeitä ja toimii ylimääräisenä liikkeen organellina.
Ravitsemusmenetelmän mukaan (assimilaatio) jaetaan:
Lisääntyminen on yleensä aseksuaalista, pituussuuntaisen jakautumisen kautta kahteen osaan. Monilla lajeilla on myös sukupuoliprosessi, jonka aikana sukupuoliset muodot sulautuvat yhteen (parpulaatio).
Pysähdyksissä makean veden säiliöissä elää lukuisia vihreitä bichenolajeja (euglena ja phytomonads). Toisinaan niitä esiintyy niin suuria määriä, että vesi muuttuu vihreäksi (vesikukinta).
Yksinäiset vapaasti elävät muodot
Yksittäisten muotojen tyypillinen edustaja on vihreä euglena (Euglena viridis). Sen rungon muoto on fusiform, vakio protoplasman ulkokerroksen tiivistymisen vuoksi. Sytoplasmassa, kehon takaosassa, on suuri pallomainen ydin.
Vihreän euglenan rungon etupäässä on ohut siima, jonka pyörimisen ansiosta se liikkuu. Siellä on myös sykkivä tyhjiö, sen säiliö ja useita klorofylliä sisältäviä kromatoforeja, joissa hiilihydraatteja syntetisoidaan hiilidioksidista ja vedestä (fotosynteesi) valossa, ts. havaitaan autotrofinen ravitsemus. Fotosynteesin tuotteet kerrostuvat tärkkelyksen kaltaisen aineen - paramyylin - muodossa. Euglena etsii valaistuja paikkoja valoherkän silmän avulla - punaisen värillisen fotoreseptorin avulla, joka sijaitsee myös kehon etuosassa.
Euglena voi myös ruokkia holotsoimalla tavalla nielemällä muotoiltuja ruokahiukkasia. Jos ympäristössä on runsaasti liuenneita eloperäinen aine, sitten ne tulevat euglenan kehoon osmoosin avulla. Siten euglenan rakenteessa ja ravitsemustavassa havaitaan eläimen ja kasviorganismin ominaisuuksien yhdistelmä.
Tietyn suolojen pitoisuuden ylläpitäminen sytoplasmassa, eli osmoottinen paine, suoritetaan erityisellä organoidilla - sykkivällä tai supistuvalla vakuolilla, joka sijaitsee kehon etupäässä. Sykkivän tyhjiön tilavuus kasvaa ajoittain. Saavutettuaan tietyn koon se kaataa kertyneen nesteen säiliön läpi ja alkaa hetken kuluttua kasvaa uudelleen. Neste tulee vakuoliin adduktorikanavien kautta.
Euglenan lisääntyminen on suvutonta, elimistön pitkittäisen jakautumisen kautta. Ensin tuma jakautuu, perusrunko, kromatoforit kaksinkertaistuvat ja sitten sytoplasma jakautuu. Siima hylätään tai siirtyy yhdelle yksilölle, ja toisessa se muodostuu uudelleen. Jako pituussuunnassa on yksi ominaispiirteet flagella-luokka. Epäsuotuisissa olosuhteissa esiintyy cystation: siima vetäytyy, euglena pyöristyy ja muodostaa tiheän kuoren. Euglena voi jakautua yhden tai useamman kerran jäädessään kuorien sisään.
Erityyppiset euglena-tyypit ovat ominaisia vesistöille, joilla on vaihteleva orgaanisen pilaantumisen aste, joten ne voivat yhdessä levien kanssa toimia kriteerinä vesihuoltolähteiden hygieenisessä arvioinnissa.
Siirtomaamuodot

Koloniaalisia siimaelimiä (volvox, pandorina, eudorina jne.) pidetään siirtymämuotoina yksisoluisista monisoluisiin organismeihin. Yksinkertaisimmin järjestetyt pesäkkeet koostuvat 4-16 täysin identtisestä yksisoluisesta yksilöstä, jotka liittyvät toisiinsa - zooideihin. Jokaisessa zooidissa on siima, silmäsilmä, kromatoforit ja supistuva vakuoli.
Siirtomaasiimalajin edustaja - volvox (Volvox globator) muodostaa suuria pallomaisia pesäkkeitä, jotka koostuvat useista tuhansista vegetatiivisista zooideista - pienistä päärynänmuotoisista soluista, joista jokaisessa on kaksi siimat. Pallon halkaisija on 1-2 mm. Sen ontelo on täytetty hyytelömäisellä aineella. Kaikki Volvox-solut (eläimet) on yhdistetty toisiinsa ohuilla protoplasmisilla silloilla, mikä mahdollistaa siipien liikkeen koordinoinnin. Pesäke liikkuu vedessä yksittäisten yksilöiden lippujen koordinoidun liikkeen vuoksi.
Volvoxissa havaitaan jo pesäkkeen solujen toiminnan jakautuminen. Joten pesäkkeen yhdessä navassa, jota pitkin se liikkuu eteenpäin, on soluja, joilla on kehittyneemmät valoherkät silmät, ja pesäkkeen alaosassa (jossa silmät ovat heikosti kehittyneet) on soluja, jotka kykenevät jakautumaan (lisääntymiseen). solut, generatiiviset zooidit), so. havaitaan erottelu somaattisiksi ja seksuaalisiksi yksilöiksi.
Volvoxin lisääntyminen tapahtuu erityisten - generatiivisten - zooidien kustannuksella. Ne jättävät pinnan pesäkkeiden sisään ja muodostavat täällä jakautumalla kertomalla tytäryhdyskuntia. Äitiyhdyskunnan kuoleman jälkeen tytärpojat aloittavat itsenäisen elämän. Syksyllä generatiivisten yksilöiden ansiosta muodostuu myös sukupuolimuotoja: suuria liikkumattomia makrogameetteja (naaraspuoliset lisääntymiszoidit) ja pienet kahdella narulla varustettuja mikrogameetit (urossyntyiset zooidit). Gametogeneesin aikana makrogameeteiksi muuttuvat yksilöt eivät jakautu ja lisää kokoa. Mikrogameetteja tuottavat yksilöt jakautuvat toistuvasti ja muodostuvat iso luku pieniä biflagellate yksilöitä. Mikrogameetit etsivät aktiivisesti liikkumattomia makrogameetteja ja sulautuvat niihin muodostaen tsygootteja. Tsygootit synnyttävät uusia pesäkkeitä. Tsygootin kaksi ensimmäistä jakautumista ovat meioottisia. Näin ollen siirtomaa-siimolla vain tsygootilla on diploidinen kromosomisarja, kaikki muut elinkaaren vaiheet ovat haploideja.
Tilaa ensisijaiset monadit (Protomonadina) Trypanosomit (Trypanosomatidae) Leishmania-suku (Leishmania)
Leishmania - leishmaniaasin aiheuttajat - tarttuvat taudit luonnollisilla pesäkkeillä. Leishmaniat löysi venäläinen lääkäri P. F. Borovsky vuonna 1898. Leishmania-suvun edustajat, jotka kuuluvat Trypanosome-perheeseen, ovat tärkeimpiä, joiden erottava piirre on kyky muodostaa useita morfologisesti. useita muotoja riippuen olemassaolon ehdoista. Muodonmuutosta esiintyy sekä selkärangattomissa että selkärankaisissa isännissä.


Siellä on seuraavat morfologisia muotoja:
- trypanosomaalinen muoto - jolle on ominaista litistetty nauhamainen runko, jonka keskellä on soikea ydin. Siima alkaa ytimen takaa. Siiman aksiaalinen filamentti menee kehon etupäähän muodostaen hyvin kehittyneen aaltoilevan kalvon. Vartalon etupäässä se päättyy, ja flagellum työntyy eteenpäin muodostaen pitkän vapaan pään.
- Kriittinen muoto - siima alkaa hieman ytimen etupuolelta, liikkuu eteenpäin muodostaen lyhyen aaltoilevan kalvon ja vapaan pään.
- leptomonas -muoto - siima alkaa vartalon etupään aivan reunasta, aaltoilevaa kalvoa ei ole, siimapään vapaa pää on huomattavan pitkä.
- leishmaniaalinen muoto - sillä on pyöreä muoto ja suuri pyöreä ydin. Sauvan muotoinen kinetoplasti sijaitsee kehon etupäässä. Siima joko puuttuu tai on vain sen solunsisäinen osa, se ei mene kehon ulkopuolelle.
- metasyklinen muoto - samanlainen kuin kriittinen, mutta vailla vapaata siimapäätä. (Kuva 1).
Leishmania-suvun lipuilla on kaksi morfologista muotoa - leptomonas ja leishmaniaalinen (kuva 2) tai solunsisäinen.
Leishmaniat jaetaan dermatotrooppisiin (sijaitsee ihossa) ja viscerotrooppisiin lajeihin (sijaitsevat sisäelimissä).
Viskeraalisen leishmaniaasin (Leischmania donovani, L. infantum) aiheuttaja
Lokalisointi. Maksan, pernan, luuytimen, imusolmukkeiden, ihonalaisen kudoksen retikuloendoteliaaliset solut.
. Viskeraalinen leishmaniaasi on yleisin Intiassa (patogeeni L. donovani), sitä esiintyy Välimeren maissa, Transkaukasiassa ja Keski-Aasiassa (patogeeni L. infantum), missä sitä kutsutaan kala-azariksi. Useissa maissa Aasiassa, Afrikassa ja Etelä-Amerikka Leishmaniaasisairauksia aiheuttavat myös muut leishmaniatyypit.
. Leptomonad ja Leishmanial muodot.

SISÄÄN Viime aikoina Leishmanian on myös havaittu asuvan ihon retikuloendoteliaalisissa soluissa, mikä selittää hyttysten tartuntatavan. Vaikuttavat solut muodostavat joskus jatkuvan kerroksen tai keskittyvät lähelle hikirauhasia ja verisuonia.
Patogeeninen toiminta. On epäsäännöllistä, jatkuvaa kuumetta. Perna ja maksa kasvavat vähitellen ja voivat saavuttaa valtavia kokoja (kuva 3). Uupumus kehittyy, punasolujen pitoisuus veressä laskee, esiintyy anemiaa. Sairaus voi olla akuutti tai krooninen (1-3 vuotta). Kuolleisuus on erittäin korkea. Useimmiten lapset ovat sairaita.
Ennaltaehkäisy: henkilökohtainen - henkilökohtainen suoja hyttysten pistoja vastaan; julkinen - joukko toimenpiteitä hyttysten ja jyrsijöiden torjumiseksi, kulkukoirien ja leishmaniaasikoirien, sakaalien tuhoamiseksi. Samaan aikaan on tarpeen suorittaa sanitaar-kasvatustyötä ja potilaiden hoitoa.
Ihon leishmaniaasi (Leishmania tropica)
Leishmanian dermatotrooppisista lajeista tunnetaan kolme alalajia: L. tropica minor ja L. tropica major (itäisellä pallonpuoliskolla) ja L. tropica mexicana (läntisellä pallonpuoliskolla).
Maantieteellinen levinneisyys. Levinnyt useissa maissa Euroopassa, Aasiassa, Amerikassa ja Afrikassa. Venäjällä L. tropican (Borovskyn taudin) aiheuttama iholeishmaniaasi on hävitetty, kun taas L. majoria esiintyy Transkaukasuksella ja Keski-Aasiassa, erityisesti useilla Turkmenistanin alueilla.
Morfofysiologiset ominaisuudet. Leptomonas ja leishmaniaaliset muodot ovat erottamattomia viscerotrooppisen leishmanian muodoista.
Elinkaari. Lähes ei eroa viskeraalisen leishmaniaasin aiheuttajasta. Tartunnan lähde ovat ihmiset ja luonnonvaraiset eläimet (hiekkaisten puoliaavioiden ja aavikoiden vyöhykkeillä elävät pienet jyrsijät - gerbiilit, maa-oravat, hamsterit, tietyntyyppiset rotat ja hiiret). Säiliöeläinten tartunta luonnollisissa olosuhteissa saavuttaa joskus 70%. Eläinten sairaus ilmenee myös ihohaavoina. Hyttyset toimivat kantajina. Säiliön jyrsijät ja vektorihyttyset ovat läheisessä kosketuksessa. Jyrsijäpesä on pysyvä elin- ja kasvualusta hyttysille, joiden tartunta voi nousta 35 %:iin. Ihmisen rooli iholeishmaniaasin leviämisessä on pieni, lukuun ottamatta joitakin maapallon alueita (Intia).
Patogeeninen toiminta. Ihon leishmaniaasissa iholle muodostuu pyöreitä, pitkiä (noin vuoden) ei-parantuvia haavaumia. avoimet osat vartalolle, erityisesti kasvoille. Paranemisen jälkeen jäljelle jää vääristävä arpi (kuva 4). Sairauden jälkeen immuniteetti kestää eliniän.
Laboratoriodiagnostiikka. Vuotohaavojen mikroskooppinen tutkimus.
Ennaltaehkäisy: henkilökohtainen - henkilökohtainen suoja hyttysten pistoja vastaan; on suositeltavaa antaa profylaktiset rokotukset eläimen iholeishmaniaasikantoja vastaan suljetuille ihoalueille. Tällaiset rokotukset suojaavat edelleen vääristäviä haavaumia ja arpia vastaan kasvoissa ja muissa paljaissa kehon osissa. Julkisena ehkäisynä torjutaan hyttysiä ja jyrsijöitä. Erityisesti he vievät kloropikriinia koloihin, kyntävät gerbiilien asuttamia maa-alueita kylien viereisillä alueilla.

Suku Trypanosoma
Trypanosoma gambiense on trypanosomiaasin, afrikkalaisen unihäiriön, aiheuttaja ihmisillä.
Lokalisointi. Ihmisten ja muiden selkärankaisten kehossa se elää veriplasmassa, imusolmukkeissa, imusolmukkeissa, aivo-selkäydinnesteessä, selkäytimen kudoksissa ja aivoissa.
Maantieteellinen levinneisyys. Löytyy päiväntasaajan Afrikasta. Taudin pesäkkeissä antilooppien verestä löydettiin trypanosomi, joka toimii sen luonnollisena säiliönä. Kantaja on afrikkalainen verta imevä hyönteinen - tse-tse-kärpäs (Glossina palpalis). Tautia esiintyy vain tämän kärpäsen levinneisyysalueella.
Morfofysiologiset ominaisuudet. Koko 13-39 mikronia. Runko on kaareva, litistetty yhteen tasoon, molemmista päistä kaventunut, varustettu yhdellä flagellulla ja aaltoilevalla kalvolla. Ruokkii osmoottisesti. Lisääntyminen tapahtuu aseksuaalisesti, pituussuuntaisen jakautumisen kautta.
Elinkaari. T. gambiensen aiheuttama trypanosomiaasi on tyypillinen tarttuva tauti, jolla on luonnollisia pesäkkeitä. Trypanosomiaasin aiheuttaja kehittyy isäntien vaihtuessa. Ensimmäinen osa trypanosomin elinkaaresta tapahtuu tsetse-kärpäsen (Glossina palpalis) ruoansulatuskanavassa, jonka alueen ulkopuolella trypanosomiaasi (afrikkalainen univitsaus) ei esiinny, toinen osa - selkärankaisten (nautaeläinten) kehossa. ja pienet nautakarjat, siat, koirat, jotkut luonnonvaraiset eläimet) sekä ihmiset. Kun kärpänen imee sairaan ihmisen verta, trypanosomit pääsevät sen vatsaan. Täällä ne lisääntyvät ja käyvät läpi useita vaiheita. Koko kehityssykli on noin 20 päivää. Kärpäset, joiden kehossa on invasiivisen vaiheen trypanosomeja, voivat tartuttaa ihmistä purettaessa.
Patogeeninen toiminta. Potilaalla, jolla on trypanosomiaasi, on vakavia keskushermoston vaurioita hermosto: havaittu lihasheikkous, uupumus, henkinen masennus, uneliaisuus. Sairaustila kestää 7-10 vuotta, ja jos sitä ei hoideta, se päättyy kuolemaan.
Laboratoriodiagnostiikka. Laboratoriodiagnostiikkaa varten tutkitaan veri, imusolmukkeiden pisteet ja aivo-selkäydinneste.
Ennaltaehkäisy. Henkilökohtainen ennaltaehkäisy tarkoittaa lääkkeiden ottamista, jotka voivat suojata tartunnalta tsetse-kärpäsen puremana. Julkinen ennaltaehkäisy koostuu kantajan tuhoamisesta. Tätä tarkoitusta varten kärpästen lisääntymispaikat - pensaat asunnon lähellä ja vesistöjen rannoilla - käsitellään hyönteismyrkkyillä tai leikataan pois.
Tilaa polyflagellaatteja (Polymastigina)

Suoliston trichomonas (Trichomonas hominis). Aiheuttaa suoliston trikomoniaasia.
Lokalisointi. Kaksoispiste.
Maantieteellinen levinneisyys. Joka paikassa.
Morfofysiologiset ominaisuudet. Runko on muodoltaan soikea, jonka takapäässä on terävä kasvusto ja yksi kuplan muotoinen ydin. Vartalon pituus on 5-15 mikronia, 4 vapaata siipiä ulottuu etupäästä eteenpäin ja yksi taaksepäin suunnattu, joka on yhteydessä aaltoilevaan kalvoon. Runko on lävistetty keskeltä tukitankolla, joka päättyy rungon takapäässä olevaan terävään piikkiin. Solun suu sijaitsee lähellä ydintä. Sytoplasmassa on ruoansulatusvakuoleja, jotka sulattavat bakteereja ja solun suun kautta nieltyjä suoliston sisältöä. Osmoottinen ravinto on myös mahdollista - nestemäiset aineet. Lisääntyminen on aseksuaalista, pitkittäisjakauman kautta. Kystien muodostuskyky on kiistanalainen.
Infektio tapahtuu vihannesten ja hedelmien, likaisten käsien kautta, jotka ovat saastuttaneet Trichomonas-pitoisia ulosteita, sekä keittämättömän veden kautta.
Patogeeninen toiminta ei ole todistettu, koska suoliston Trichomonas esiintyy terveiden ihmisten ja potilaiden suolistossa. On olemassa mielipide, että suoliston trichomonas ei aiheuta tautia, vaan se liittyy vain muiden syiden aiheuttamiin patologisiin prosesseihin.
Laboratoriodiagnostiikka. Ulosteen sivelysolujen mikroskooppinen tutkimus, vegetatiivisten muotojen löytäminen ulosteista.
Ennaltaehkäisy. Henkilökohtainen käsien pesu ennen ruokailua ja vessassa käynnin jälkeen, ruoan ja juomaveden lämpökäsittely, raakana syötyjen vihannesten ja hedelmien perusteellinen pesu, ruoan ja veden suojaaminen pölyltä ja kärpäsiltä, jotka voivat olla mekaanisia eri taudinaiheuttajien kantajia. Julkinen ennaltaehkäisy koostuu vesihuollon, elintarvikeyritysten ja päivittäistavarakauppojen, julkisten paikkojen saniteettitilan seurannasta, kärpästen torjunnasta ja hygieniatiedon levittämisestä.

Virtsa- ja sukuelinten trichomonas (Trichomonas vaginalis)- urogenitaalisen trikomonadoosin aiheuttaja.
Lokalisointi. Miesten ja naisten virtsatie.
Maantieteellinen levinneisyys. Joka paikassa.
Morfofysiologiset ominaisuudet. Rakenne on hyvin samanlainen kuin suoliston Trichomonas. Erottavia piirteitä ovat rungon suuret koot (pituus vaihtelee 7-30 mikronia). Vartalon muoto on useimmiten päärynän muotoinen, siinä on 4 lippua, aaltoileva kalvo ja piikkiin päättyvä tukitanko.
Patogeeninen toiminta. Aiheuttaa sukupuolielinten tulehduksellisia prosesseja, jotka ovat pitkittyneet. Uskotaan, että altistava hetki patogeenisyyden ilmenemiselle on tietyntyyppisten bakteerien esiintyminen sukuelinten alueella.
Naisilla emätin vaikuttaa aluksi, mutta myöhemmin sairaus muuttuu multifokaaliseksi. Akuutissa taudissa on ominaista runsas nestevuoto, kutina ja polttaminen. Miehillä suurin osa taudista on oireetonta.
Infektio tapahtuu seksuaalisesti sekä käytettäessä vuodevaatteita, liinavaatteita, potilaan sientä. Infektio on mahdollinen, kun gynekologi tutkii saastuneiden instrumenttien ja käsineiden kautta.
Laboratoriodiagnostiikka. Virtsateiden eritteiden mikroskooppinen tutkimus. Diagnoosi tehdään potilaan eritteistä löytyneiden vegetatiivisten muotojen perusteella.
Ennaltaehkäisy. määräytyy tartuntatavan mukaan.

Lamblia (Lamblia intestinalis). Aiheuttaa taudin giardiaasia. Tämän flagellaatin löysi professori Harkovan yliopisto D. F. Lamblem vuonna 1859
Lokalisointi. Pohjukaissuoli voi toisen kerran tunkeutua sappitiehyisiin.
Maantieteellinen levinneisyys. Joka paikassa.
Laboratoriodiagnostiikka. Kystien havaitseminen ulosteissa tai vegetatiivisissa muodoissa pohjukaissuolen sisällöstä koettimen aikana.
Ennaltaehkäisy. a) elintarvikkeiden ja juomaveden lämpökäsittely; kiehuminen tappaa giardiakystat; b) hedelmien, vihannesten ja muiden raakana kulutettujen ruokien perusteellinen pesu; c) keitetyn ruoan ja juomaveden säilytys suljetussa astiassa. On pidettävä mielessä, että Giardia-kystat voivat joutua pölyn kanssa ruoalle ja kärpästen kautta; d) taistelu kärpäsiä vastaan. Kärpäset vierailevat jäteveden kaatopaikoilla, jätealtaissa, ne istuvat ihmisten ulosteiden päällä, mätänevän ruoan päällä ja värjättyään ruumiinsa ulosteissa lentävät ihmisen asuntoon, missä ne istuvat ruoan, astioiden, ihmisen ihon päällä; e) käsien pesu ennen ateriaa ja wc-käynnin jälkeen.




