Mitä molekyylibiologia tutkii. Molekyylibiologia - Molekyylibiologia. Perinnöllisyyden molekyyliperusta
Molekyylibiologia koki omien tutkimusmenetelmiensä nopean kehityksen ajanjakson, joka nyt eroaa biokemiasta. Näitä ovat erityisesti geenitekniikan menetelmät, kloonaus, keinotekoinen ilmentäminen ja geenin poisto. Koska DNA on geneettisen tiedon aineellinen kantaja, molekyylibiologia on tullut paljon lähemmäksi genetiikkaa, ja risteyksessä muodostui molekyyligenetiikka, joka on sekä genetiikan että molekyylibiologian osa. Aivan kuten molekyylibiologia käyttää laajasti viruksia tutkimusvälineenä, virologia käyttää molekyylibiologian menetelmiä ongelmiensa ratkaisemiseen. Analyysia varten geneettistä tietoa mukana tietotekniikka, jonka yhteydessä on ilmaantunut uusia suuntauksia molekyyligenetiikka joita joskus pidetään erillisinä tieteenaloina: bioinformatiikka, genomiikka ja proteomiikka.
Kehityksen historia
Tämä tärkeä löytö valmisteltiin pitkällä virusten ja bakteerien genetiikan ja biokemian tutkimusvaiheella.
Vuonna 1928 Frederick Griffith osoitti ensimmäisen kerran, että lämmöllä tapettujen patogeenisten bakteerien uute voi siirtää patogeenisyyden hyvänlaatuisiin bakteereihin. Bakteeritransformaatioiden tutkiminen johti edelleen taudinaiheuttajan puhdistukseen, joka vastoin odotuksia osoittautui ei proteiiniksi, vaan nukleiinihapoksi. Nukleiinihappo itsessään ei ole vaarallinen, se kantaa vain geenejä, jotka määräävät mikro-organismin patogeenisuuden ja muut ominaisuudet.
XX-luvun 50-luvulla osoitettiin, että bakteereilla on primitiivinen seksuaalinen prosessi, ne pystyvät vaihtamaan ekstrakromosomaalista DNA:ta, plasmideja. Plasmidien löytäminen sekä transformaatiot muodostivat perustan molekyylibiologiassa yleiselle plasmiditeknologialle. Toinen menetelmälle tärkeä löytö oli 1900-luvun alussa löydetty bakteerivirukset, bakteriofagit. Faagit voivat myös siirtää geneettistä materiaalia yhdestä bakteerisolu toiselle. Faagien aiheuttama bakteeritartunta johtaa muutokseen bakteeri-RNA:n koostumuksessa. Jos RNA:n koostumus on ilman faageja samanlainen kuin bakteeri-DNA:n koostumus, niin infektion jälkeen RNA:sta tulee samankaltaisempi kuin bakteriofagi-DNA. Siten havaittiin, että RNA:n rakenne määräytyy DNA:n rakenteen perusteella. Proteiinisynteesin nopeus soluissa puolestaan riippuu RNA-proteiinikompleksien määrästä. Näin se muotoiltiin molekyylibiologian keskeinen dogma: DNA ↔ RNA → proteiini.
Molekyylibiologian jatkokehitystä seurasi sekä sen metodologian kehitys, erityisesti menetelmän keksiminen DNA:n nukleotidisekvenssin määrittämiseksi (W. Gilbert ja F. Sanger, Nobelin kemianpalkinto 1980), että uusia löydöt geenien rakenteen ja toiminnan tutkimuksen alalla (ks. Genetiikan historia). TO XXI alku luvulla saatiin tietoa ihmisen kaiken DNA:n ja useiden muiden lääketieteen, maatalouden ja tieteellisen tutkimuksen kannalta tärkeimpien organismien primäärirakenteesta, mikä johti useiden uusien biologian alueiden syntymiseen: genomiikka, bioinformatiikka jne.
Katso myös
- Molekyylibiologia (lehti)
- Transkriptomiikka
- Molekyylipaleontologia
- EMBO - Euroopan järjestö molekyylibiologit
Kirjallisuus
- Laulaja M., Berg P. Geenit ja genomit. - Moskova, 1998.
- Stent G., Kalindar R. Molekyyligenetiikka. - Moskova, 1981.
- Sambrook J., Fritsch E.F., Maniatis T. Molekyylikloonaus. - 1989.
- Patrushev L.I. Geenien ilmentyminen. - M.: Nauka, 2000. - 000 s., ill. ISBN 5-02-001890-2
Linkit
Wikimedia Foundation. 2010 .
- Ardatovsky-alue Nižni Novgorodin alueella
- Arzamasin alue Nižni Novgorodin alueella
Katso, mitä "Molecular Biology" on muissa sanakirjoissa:
MOLEKYYLIBIOLOGIA- opiskelee perusteita. elämän ominaisuudet ja ilmentymät molekyylitasolla. Tärkeimmät suunnat M. b. ovat rakenteellisia tutkimuksia toimiva organisaatio solujen geneettinen laite ja perinnöllisen tiedon toteuttamismekanismi ... ... Biologinen tietosanakirja
MOLEKYYLIBIOLOGIA- tutkii elämän perusominaisuuksia ja ilmenemismuotoja molekyylitasolla. Selvittää, miten ja missä määrin organismien kasvu ja kehitys, perinnöllisen tiedon varastointi ja välittäminen, energian muuntaminen elävissä soluissa ja muut ilmiöt johtuvat ... Suuri Ensyklopedinen sanakirja
MOLEKYYLIBIOLOGIA Nykyaikainen tietosanakirja
MOLEKYYLIBIOLOGIA- MOLEKULAARIBIOLOGIA, biologinen tutkimus elävien organismien muodostavien MOLEKYELIEN rakenteesta ja toiminnasta. Pääasialliset opiskelualueet ovat fyysinen ja Kemialliset ominaisuudet proteiinit ja nukleiinihapot, kuten DNA. Katso myös… … Tieteellinen ja tekninen tietosanakirja
molekyylibiologia- Biol.:n osa, joka tutkii elämän perusominaisuuksia ja ilmenemismuotoja molekyylitasolla. Selvittää, miten ja missä määrin organismien kasvu ja kehitys, perinnöllisen tiedon varastointi ja välittäminen, energian muuntaminen elävissä soluissa ja ... ... Mikrobiologian sanakirja
molekyylibiologia- — Biotekniikan aiheet FI molekyylibiologia… Teknisen kääntäjän käsikirja
Molekyylibiologia- MOLEKULAARIBIOLOGIA, tutkii elämän perusominaisuuksia ja ilmenemismuotoja molekyylitasolla. Selvittää, miten ja missä määrin organismien kasvu ja kehitys, perinnöllisen tiedon varastointi ja välittäminen, energian muuntaminen elävissä soluissa ja ... ... Kuvitettu tietosanakirja
Molekyylibiologia- tiede, joka asettaa tehtäväkseen elämänilmiöiden luonteen tuntemisen tutkimalla biologisia esineitä ja järjestelmiä molekyylitasoa lähestyvällä ja joissakin tapauksissa tämän rajan saavuttamisessa. Tämän lopullinen tavoite on…… Suuri Neuvostoliiton tietosanakirja
MOLEKYYLIBIOLOGIA- tutkii elämänilmiöitä makromolekyylien tasolla (ch. arr. proteiinit ja nukleiinihapot) soluttomissa rakenteissa (ribosomit jne.), viruksissa ja myös soluissa. M:n tarkoitus. näiden makromolekyylien roolin ja toimintamekanismin määrittäminen perustuen ... ... Chemical Encyclopedia
molekyylibiologia- tutkii elämän perusominaisuuksia ja ilmenemismuotoja molekyylitasolla. Selvittää, miten ja missä määrin organismien kasvu ja kehitys, perinnöllisen tiedon varastointi ja välittäminen, energian muuntaminen elävissä soluissa ja muut ilmiöt ... ... tietosanakirja
Kirjat
- Solun molekyylibiologia. Problem Book, J. Wilson, T. Hunt. Amerikkalaisten kirjailijoiden kirja on B. Albertsin, D. Brayn, J. Lewisin ym. oppikirjan "Molecular Biology of the Cell" liite 2. painokseen, joka sisältää kysymyksiä ja tehtäviä, joiden tarkoituksena on syventää . ..
Voidaan sanoa, että molekyylibiologia tutkii elämän ilmenemismuotoja elottomissa rakenteissa tai järjestelmissä, joissa on alkeellisia merkkejä vitaalitoiminnasta (jotka voivat olla yksittäisiä biologisia makromolekyylejä, niiden komplekseja tai organelleja), tutkien, kuinka elävää ainetta kuvaavat keskeiset prosessit toteutuvat kemiallisesti. vuorovaikutuksia ja muunnoksia.
Molekyylibiologian erottaminen biokemiasta itsenäiseksi tieteenalaksi sanelee se, että sen päätehtävänä on tutkia eri prosesseissa mukana olevien biologisten makromolekyylien rakennetta ja ominaisuuksia, selvittää niiden vuorovaikutuksen mekanismeja. Biokemia puolestaan käsittelee varsinaisten elintärkeän toiminnan prosessien, niiden etenemismallien ja näiden prosessien mukana tulevien molekyylien muunnosten tutkimusta elävässä organismissa. Lopulta molekyylibiologia yrittää vastata kysymykseen, miksi tämä tai tuo prosessi tapahtuu, kun taas biokemia vastaa kysymyksiin, missä ja miten kemian näkökulmasta kyseinen prosessi tapahtuu.
Tarina
Molekyylibiologia erillisenä biokemian osa-alueena alkoi muotoutua 1930-luvulla. Silloin elämän ilmiön syvemmälle ymmärtämiselle syntyi tarve kohdennetuille tutkimuksille molekyylitasolla elävien organismien perinnöllisen tiedon varastointi- ja siirtoprosesseista. Sitten määritettiin molekyylibiologian tehtävä rakenteen, ominaisuuksien ja vuorovaikutuksen tutkimuksessa nukleiinihapot ja proteiineja. Termiä "molekyylibiologia" käytti ensimmäisenä englantilainen tiedemies William Astbury tutkimuksessa, joka liittyi fibrillaaristen proteiinien, kuten kollageenin, veren fibriinin tai lihasten supistumisproteiinien molekyylirakenteen ja fysikaalisten ja biologisten ominaisuuksien välisen yhteyden selvittämiseen. .
Molekyylibiologian alkuaikoina RNA:ta pidettiin kasvien ja sienten komponenttina, kun taas DNA:ta pidettiin tyypillisenä eläinsolujen komponenttina. Ensimmäinen tutkija, joka osoitti, että DNA:ta löytyy kasveista, oli Andrey Nikolaevich Belozersky, joka eristi herneen DNA:n vuonna 1935. Tämä löytö vahvisti sen tosiasian, että DNA on universaali nukleiinihappo, jota esiintyy kasvi- ja eläinsoluissa.
Suuri saavutus oli George Beadlen ja Edward Tatumin selvittäminen suorasta syy-yhteydestä geenien ja proteiinien välillä. Kokeissaan he paljastivat neurosporisoluja ( Neurosporacrassa) Röntgensäteilyaltistus, joka aiheutti mutaatioita. Saadut tulokset osoittivat, että tämä johti muutokseen tiettyjen entsyymien ominaisuuksissa.
Vuonna 1940 Albert Claude eristi sytoplasmista RNA:ta sisältäviä rakeita eläinsolujen sytoplasmasta, jotka olivat pienempiä kuin mitokondriot. Hän kutsui niitä mikrosomeiksi. Myöhemmin, tutkittaessa eristettyjen hiukkasten rakennetta ja ominaisuuksia, todettiin niiden perustavanlaatuinen rooli proteiinien biosynteesiprosessissa. Vuonna 1958 ensimmäisessä näille hiukkasille omistetussa symposiumissa näitä hiukkasia päätettiin kutsua ribosomeiksi.
Toinen tärkeä askel molekyylibiologian kehityksessä oli Oswald Averyn, Colin MacLeodin ja MacLean McCarthyn vuonna 1944 tekemän kokeen julkaistut tiedot, jotka osoittivat DNA:n olevan syy bakteerien transformaatioon. Tämä oli ensimmäinen kokeellinen todiste DNA:n roolista perinnöllisen tiedon välittämisessä, mikä kumosi aiemman ajatuksen geenien proteiiniluonteesta.
Frederick Sanger osoitti 1950-luvun alussa, että proteiiniketju on ainutlaatuinen aminohappotähteiden sekvenssi. 1950-luvun lopulla Max Perutz ja John Kendrew selvittivät ensimmäisten proteiinien spatiaalisen rakenteen. Jo vuonna 2000 tunnettiin satoja tuhansia luonnollisia aminohapposekvenssejä ja tuhansia proteiinien spatiaalisia rakenteita.
Samoihin aikoihin Erwin Chargaffin tutkimus antoi hänelle mahdollisuuden muotoilla säännöt, jotka kuvaavat typpipitoisten emästen suhdetta DNA:ssa (säännöissä sanotaan, että DNA:n lajieroista riippumatta guaniinin määrä on yhtä suuri kuin sytosiinin määrä ja adeniinin määrä on sama kuin theminin määrä), joka myöhemmin auttoi tekemään suurimman läpimurron molekyylibiologiassa ja yhden suurimmat löydöt biologiassa yleensä.
Tämä tapahtuma tapahtui vuonna 1953, kun James Watson ja Francis Crick perustuivat Rosalind Franklinin ja Maurice Wilkinsin työhön. Röntgendiffraktioanalyysi DNA loi DNA-molekyylin kaksijuosteisen rakenteen. Tämä löytö teki mahdolliseksi vastata peruskysymykseen perinnöllisen tiedon kantajan kyvystä lisääntyä itse ja ymmärtää tällaisen tiedon välitysmekanismia. Samat tutkijat muotoilivat typpipitoisten emästen komplementaarisuuden periaatteen, joka on avainasemassa supramolekyylisten rakenteiden muodostumismekanismin ymmärtämisessä. Tämä periaate, jota käytetään nyt kuvaamaan kaikkia molekyylikomplekseja, mahdollistaa olosuhteiden kuvaamisen ja ennustamisen heikkojen (ei-valenttien) molekyylien välisten vuorovaikutusten syntymiselle, jotka määräävät sekundääristen, tertiääristen jne. makromolekyylien rakenteet, supramolekyylien itsekokoonpano biologiset järjestelmät jotka määrittävät niin laajan valikoiman molekyylirakenteita ja niiden toiminnallisia joukkoja. Sitten, vuonna 1953, syntyi Tiede-lehti Journal of Molecular Biology. Sitä johti John Kendrew, tieteellisiä etuja joka oli globulaaristen proteiinien rakenteen tutkimus (Nobel-palkinto vuonna 1962, yhdessä Max Perutzin kanssa). V. A. Engelhardt perusti Neuvostoliitossa samanlaisen venäjänkielisen lehden nimeltä Molecular Biology vuonna 1966.
Vuonna 1958 Francis Crick muotoili ns. molekyylibiologian keskeinen dogma: ajatus geneettisen tiedon virtauksen peruuttamattomuudesta DNA:sta RNA:n kautta proteiineihin kaavion mukaisesti DNA → DNA (replikaatio, DNA:n kopion luominen), DNA → RNA (transkriptio, geenien kopiointi), RNA → proteiini (translaatio, rakenneproteiineja koskevien tietojen dekoodaus). Tätä dogmaa korjattiin jonkin verran vuonna 1970 ottaen huomioon kertynyt tieto, koska käänteistranskription ilmiön löysivät itsenäisesti Howard Temin ja David Baltimore: löydettiin entsyymi - käänteiskopioija, joka vastaa käänteisen transkription toteuttamisesta - kaksijuosteisen DNA:n muodostuminen yksijuosteiselle RNA-templaatille, jota esiintyy onkogeenisissä viruksissa. On huomattava, että geneettisen tiedon tiukka välttämättömyys nukleiinihapoista proteiineihin on edelleen molekyylibiologian perusta.
Vuonna 1957 Alexander Sergeevich Spirin osoitti yhdessä Andrei Nikolajevitš Belozerskyn kanssa, että huolimatta merkittävistä eroista eri organismien DNA:n nukleotidikoostumuksessa, kokonais-RNA:n koostumus on samanlainen. Näiden tietojen perusteella he tulivat sensaatiomaiseen johtopäätökseen, että solun kokonais-RNA ei voi toimia geneettisen tiedon kantajana DNA:sta proteiineihin, koska se ei vastaa sitä koostumukseltaan. Samalla he huomasivat, että RNA:ta on pieni osa, joka vastaa nukleotidikoostumukseltaan täysin DNA:ta ja joka voi olla todellinen geneettisen tiedon kantaja DNA:sta proteiineihin. Tämän seurauksena he ennustivat suhteellisen pienten RNA-molekyylien olemassaolon, jotka ovat rakenteeltaan analogisia yksittäisten DNA-osien kanssa ja toimivat välittäjinä DNA:n sisältämän geneettisen tiedon siirtämisessä ribosomiin, jossa proteiinimolekyylejä syntetisoidaan tämän tiedon avulla. Vuonna 1961 (S. Brenner, F. Jacob, M. Meselson toisaalta ja F. Gros, Francois Jacob ja Jacques Monod olivat ensimmäiset, jotka vahvistivat kokeellisesti tällaisten molekyylien – informaatio- (matriisi) RNA:n – olemassaolon. he kehittivät DNA:n funktionaalisten yksiköiden - operonin - käsitteen ja mallin, joka mahdollisti tarkasti, kuinka geeniekspression säätely prokaryooteissa tapahtuu. rakenteellinen organisaatio ja molekyylikoneiden - ribosomien - työ mahdollisti geneettisen tiedon liikkumista kuvaavan postulaatin, jota kutsutaan molekyylibiologian keskeiseksi dogmaksi: DNA - mRNA - proteiini.
Vuonna 1961 ja muutaman seuraavan vuoden aikana Heinrich Mattei ja Marshall Nirenberg sekä sitten Har Korana ja Robert Holley suorittivat useita tutkimuksia geneettisen koodin tulkitsemiseksi, minkä seurauksena DNA-rakenteen ja syntetisoitujen proteiinien välille muodostui suora yhteys. ja nukleotidisekvenssi, joka määrittää proteiinin aminohappojoukon. Saatiin myös tietoa geneettisen koodin universaalisuudesta. Löydökset palkittiin Nobel-palkinnolla vuonna 1968.
Kehitystä varten nykyajan ideoita RNA:n toiminnoista ratkaiseva oli ei-koodaavan RNA:n löytäminen, joka tehtiin Aleksanteri Sergeevich Spirinin yhdessä Andrei Nikolajevitš Belozerskin kanssa vuonna 1958, Charles Brennerin kanssa tekijöiden ja Saul Spiegelmanin kanssa vuonna 1961. . Tämän tyyppinen RNA muodostaa suurimman osan solujen RNA:sta. Ribosomaaliset RNA:t ovat pääasiassa ei-koodaavia.
Eläinsolujen viljely- ja hybridisointimenetelmät ovat saaneet vakavaa kehitystä. Vuonna 1963 François Jacob ja Sydney Brenner muotoilivat ajatuksen replikonista, luontaisesti replikoituvien geenien sekvenssistä, joka selittää geenien replikaation säätelyn tärkeitä näkökohtia.
Vuonna 1967 A. S. Spirinin laboratoriossa osoitettiin ensimmäistä kertaa, että kompaktisti laskostuneen RNA:n muoto määrittää ribosomaalisen partikkelin morfologian.
Vuonna 1968 tehtiin merkittävä perustavanlaatuinen löytö. Okazaki, löydettyään jäljessä olevan juosteen DNA-fragmentteja replikaatioprosessin tutkimuksessa, nimesi Okazaki-fragmentit hänen mukaansa, selvensi DNA:n replikaation mekanismia.
Vuonna 1970 Howard Temin ja David Baltimore tekivät itsenäisesti merkittävän löydön: löydettiin entsyymi - käänteiskopioijaentsyymi, joka vastaa käänteistranskription toteuttamisesta - kaksijuosteisen DNA:n muodostuminen yksijuosteiselle RNA-templaatille, joka tapahtuu onkogeeniset virukset, jotka sisältävät RNA:ta.
yksi vielä tärkeä saavutus molekyylibiologia oli selitys mutaatioiden mekanismista molekyylitasolla. Tutkimussarjan tuloksena määritettiin mutaatioiden päätyypit: duplikaatiot, inversiot, deleetiot, translokaatiot ja transpositiot. Tämä mahdollisti evoluutiomuutosten tarkastelun geeniprosessien näkökulmasta ja mahdollisti fylogeniassa käytettävän molekyylikellojen teorian kehittämisen.
1970-luvun alkuun mennessä oli muotoiltu nukleiinihappojen ja proteiinien toiminnan perusperiaatteet elävässä organismissa. Todettiin, että proteiinit ja nukleiinihapot kehossa syntetisoituvat matriisimekanismin mukaisesti, matriisimolekyyli kuljettaa salattua tietoa aminohappojen (proteiinissa) tai nukleotidien (nukleiinihapossa) sekvenssistä. Replikaation (DNA:n kaksinkertaistuminen) tai transkription (mRNA:n synteesi) aikana DNA toimii sellaisena matriisina, translaation (proteiinisynteesi) tai käänteistranskription aikana - mRNA.
Siten luotiin teoreettiset edellytykset molekyylibiologian soveltavien alojen, erityisesti geenitekniikan, kehittämiselle. Vuonna 1972 Paul Berg, Herbert Bauer ja Stanley Cohen kehittivät molekyylikloonausteknologiaa. Sitten he olivat ensimmäiset, jotka saivat rekombinantti-DNA:n in vitro. Nämä erinomaiset kokeet loivat perustan geenitekniikalle, ja tätä vuotta pidetään tämän tieteellisen suunnan syntymäpäivänä.
Vuonna 1977 Frederick Sanger ja itsenäisesti Allan Maxum ja Walter Gilbert kehittivät erilaisia menetelmiä ensisijainen rakenne(sekvensointi) DNA. Sanger-menetelmä, ns. ketjun lopetusmenetelmä, on nykyaikaisen sekvensointimenetelmän perusta. Sekvensoinnin periaate perustuu leimattujen emästen käyttöön, jotka toimivat terminaattoreina syklisessä sekvensointireaktiossa. Tästä menetelmästä on tullut laajalle levinnyt, koska analyysi on nopeaa.
1976 - Frederick. Sanger selvitti 5375 nukleotidiparin pituisen faagin φΧ174 DNA:n nukleotidisekvenssin.
1981 - Sirppisoluanemiasta tulee ensimmäinen geneettinen sairaus, joka diagnosoidaan DNA-testauksella.
1982-1983 RNA:n katalyyttisen toiminnan löytö T. Checkin ja S. Altmanin amerikkalaisissa laboratorioissa muutti olemassa olevia käsityksiä proteiinien yksinomaisesta roolista. Analogisesti katalyyttisten proteiinien - entsyymien kanssa, katalyyttisiä RNA:ita kutsuttiin ribotsyymeiksi.
1987 Keri Mullez löysi polymeraasiketjureaktion, jonka ansiosta on mahdollista lisätä keinotekoisesti merkittävästi DNA-molekyylien määrää liuoksessa jatkotyötä varten. Nykyään se on yksi tärkeimmistä molekyylibiologian menetelmistä, joita käytetään perinnöllisten ja virussairauksien tutkimuksessa, geenien tutkimuksessa sekä geneettisessä tunnistamisessa ja sukulaisuussuhteessa jne.
Vuonna 1990 samaan aikaan kolme tutkijaryhmää julkaisi menetelmän, jonka avulla saatiin nopeasti synteettisiä toiminnallisesti aktiivisia RNA:ita laboratoriossa (keinotekoisia ribotsyymejä tai molekyylejä, jotka ovat vuorovaikutuksessa eri ligandien - aptameerien kanssa). Tätä menetelmää kutsutaan "evoluutioksi in vitro". Ja pian sen jälkeen, vuosina 1991-1993 A.B.:n laboratoriossa. Chetverinalle osoitettiin kokeellisesti mahdollisuus RNA-molekyylien olemassaoloon, kasvuun ja monistumiseen pesäkkeiden muodossa kiinteällä alustalla.
Vuonna 1998, lähes samanaikaisesti, Craig Mello ja Andrew Fire kuvasivat mekanismia, joka havaittiin aiemmin bakteereiden ja kukkien geenikokeissa. RNA:n häiriö, jossa pieni kaksijuosteinen RNA-molekyyli johtaa spesifiseen geeniekspression suppressioon.
RNA-häiriön mekanismin löytäminen on erittäin tärkeää. käytännön arvoa modernille molekyylibiologialle. Tätä ilmiötä käytetään laajasti mm tieteellisiä kokeita työkaluna "sammutukseen", toisin sanoen yksittäisten geenien ilmentymisen tukahduttamiseen. Erityisen mielenkiintoista on se, että tämä menetelmä mahdollistaa tutkittujen geenien aktiivisuuden palautuvan (tilapäisen) tukahdutuksen. Tämän ilmiön soveltamiseksi virusperäisten, neoplastisten, rappeuttavien ja aineenvaihduntasairauksien hoitoon on meneillään tutkimus. On huomattava, että vuonna 2002 löydettiin poliovirusten mutantteja, jotka voivat välttää RNA:n häiriön, joten tarvitaan lisää vaivalloista työtä todellisen tehokkaita menetelmiä tähän ilmiöön perustuva hoito.
Vuosina 1999-2001 useat tutkijaryhmät määrittelivät bakteeriribosomin rakenteen 5,5 - 2,4 angströmin resoluutiolla.
Tuote
Molekyylibiologian saavutuksia elävän luonnon tuntemisessa voidaan tuskin yliarvioida. Suuri menestys saavutettiin onnistuneen tutkimuskonseptin ansiosta: monimutkainen biologisia prosesseja tarkastellaan yksittäisten molekyylijärjestelmien näkökulmasta, mikä mahdollistaa tarkkojen fysikaalis-kemiallisten tutkimusmenetelmien käytön. Se houkutteli tälle tieteenalalle myös monia suuria mieliä läheisiltä alueilta: kemiasta, fysiikasta, sytologiasta, virologiasta, millä oli myös myönteinen vaikutus tämän alan tieteellisen tiedon laajuuteen ja kehitysnopeuteen. Sellaiset merkittävät löydöt kuin DNA:n rakenteen määrittäminen, geneettisen koodin purkaminen ja genomin keinotekoinen suunnattu muuntaminen ovat mahdollistaneet organismien kehitysprosessien erityispiirteiden ymmärtämisen paljon syvemmälle ja onnistuneesti ratkaisemaan lukuisia tärkeitä perusperiaatteita. ja sovelletut tieteelliset, lääketieteelliset ja sosiaaliset ongelmat, joita ei niin kauan sitten pidetty ratkaisemattomina.
Molekyylibiologian tutkimuskohteena ovat pääasiassa proteiinit, nukleiinihapot ja niihin perustuvat molekyylikompleksit (molekyylikoneet) ja prosesseja, joihin ne osallistuvat.
Nukleiinihapot ovat lineaarisia polymeerejä, jotka koostuvat nukleotidiyksiköistä (viisijäsenisen sokerin yhdisteistä, joissa on fosfaattiryhmä syklin viidennessä atomissa ja yksi neljästä typpipitoisesta emäksestä), jotka on yhdistetty toisiinsa fosfaattiryhmien esterisidoksella. Siten nukleiinihappo on pentoosifosfaattipolymeeri, jossa on typpipitoisia emäksiä sivusubstituentteina. Kemiallinen koostumus RNA-ketju eroaa DNA:sta siinä, että ensimmäinen koostuu viisijäsenisestä riboosin hiilihydraattisyklistä, kun taas toinen koostuu dehydroksyloidusta riboosijohdannaisesta - deoksiriboosista. Samaan aikaan nämä molekyylit eroavat dramaattisesti avaruudessa, koska RNA on joustava yksijuosteinen molekyyli, kun taas DNA on kaksijuosteinen molekyyli.
Proteiinit ovat lineaarisia polymeerejä, jotka ovat toisiinsa liittyneitä alfa-aminohappoketjuja. peptidisidos, josta heidän toinen niminsä - polypeptidit. Luonnollisten proteiinien koostumus sisältää monia erilaisia aminohappoyksiköitä - ihmisillä jopa 20 -, mikä määrää näiden molekyylien laajan valikoiman toiminnallisia ominaisuuksia. Nämä tai nuo proteiinit osallistuvat lähes kaikkiin kehon prosesseihin ja suorittavat monia tehtäviä: ne toimivat solujen rakennusmateriaalina, kuljettavat aineita ja ioneja, katalysoivat kemialliset reaktiot, tämä lista on erittäin pitkä. Proteiinit muodostavat stabiileja molekyylikonformaatioita eri organisaatiotasoilla (sekundaariset ja tertiääriset rakenteet) ja molekyylikomplekseja, mikä entisestään laajentaa niiden toiminnallisuutta. Näillä molekyyleillä voi olla korkea spesifisyys tiettyjen tehtävien suorittamiseen monimutkaisen spatiaalisen pallomaisen rakenteen muodostumisen vuoksi. Laaja valikoima proteiineja varmistaa tutkijoiden jatkuvan kiinnostuksen tällaisia molekyylejä kohtaan.
Nykyaikaiset ajatukset molekyylibiologian aiheesta perustuvat yleistykseen, jonka Francis Crick esitti ensimmäisen kerran vuonna 1958 molekyylibiologian keskeisenä dogmana. Sen ydin oli väite, että geneettinen informaatio elävissä organismeissa käy läpi tiukasti määritellyt toteutusvaiheet: kopiointi DNA:sta DNA:han perinnön sisäänkäynnissä, DNA:sta RNA:han ja sitten RNA:sta proteiiniin, ja käänteinen siirtyminen ei ole mahdollista. Tämä väite piti paikkansa vain osittain, joten myöhemmin keskeinen dogma korjattiin ottaen huomioon äskettäin löydetyt tiedot.
Tällä hetkellä on olemassa useita tapoja toteuttaa geneettistä materiaalia, jotka edustavat erilaisia sekvenssejä geneettisen tiedon kolmen tyyppisen olemassaolon toteuttamiseksi: DNA, RNA ja proteiini. Yhdeksässä mahdollisessa toteutustavassa erotetaan kolme ryhmää: nämä ovat kolme yleistä muutosta (yleistä), jotka suoritetaan normaalisti useimmissa elävissä organismeissa; kolme erityistä transformaatiota (erityinen), jotka suoritetaan joissakin viruksissa tai erityisissä laboratorio-olosuhteissa; kolme tuntematonta muunnosa (tuntematon), joiden toteuttamista pidetään mahdottomaksi.
Yleisiä muunnoksia ovat seuraavat tavat toteuttaa geneettinen koodi: DNA → DNA (replikaatio), DNA → RNA (transkriptio), RNA → proteiini (translaatio).
Perinnöllisten ominaisuuksien siirtämiseksi vanhempien on siirrettävä täysimittainen DNA-molekyyli jälkeläisilleen. Prosessia, jolla alkuperäisen DNA:n tarkka kopio voidaan syntetisoida ja siten siirtää geneettistä materiaalia, kutsutaan replikaatioksi. Sen suorittavat erityiset proteiinit, jotka purkavat molekyylin (suoristavat sen osan), purkavat kaksoiskierre ja käyttämällä DNA-polymeraasia luomaan tarkka kopio alkuperäisestä DNA-molekyylistä.
Solun elämän varmistamiseksi sen on jatkuvasti viitattava DNA:n kaksoiskierteeseen upotettuun geneettiseen koodiin. Tämä molekyyli on kuitenkin liian suuri ja kömpelö käytettäväksi suorana geneettisen materiaalin lähteenä jatkuvaan proteiinisynteesiin. Siksi DNA:han upotetun tiedon toteutuksessa on välivaihe: mRNA:n synteesi, joka on pieni yksijuosteinen molekyyli, joka on komplementaarinen tiettyä proteiinia koodaavalle DNA-segmentille. Transkriptioprosessi saadaan aikaan RNA-polymeraasin ja transkriptiotekijöiden avulla. Tuloksena oleva molekyyli voidaan sitten helposti kuljettaa proteiinisynteesistä vastaavaan solun osaan - ribosomiin.
Kun RNA on saapunut ribosomiin, alkaa geneettisen tiedon toteutumisen viimeinen vaihe. Ribosomi lukee mRNA:sta geneettinen koodi kodoneiksi kutsuttuja triplettejä ja syntetisoi vastaavan proteiinin vastaanotetun tiedon perusteella.
Erityisten transformaatioiden aikana geneettinen koodi toteutuu kaavion mukaisesti RNA → RNA (replikaatio), RNA → DNA (käänteistranskriptio), DNA → proteiini (suora translaatio). Tämän tyyppinen replikaatio toteutuu monissa viruksissa, joissa sen suorittaa RNA-riippuvainen RNA-polymeraasientsyymi. Samanlaisia entsyymejä löytyy myös eukaryoottisoluista, joissa ne liittyvät RNA:n hiljennysprosessiin. Käänteistranskriptiota on löydetty retroviruksista, joissa sen suorittaa käänteiskopioijaentsyymi, ja joissakin tapauksissa eukaryoottisoluissa, esimerkiksi telomeerisynteesin aikana. Suora lähetys tapahtuu vain keinotekoisissa olosuhteissa eristetyssä järjestelmässä solun ulkopuolella.
Mitä tahansa kolmesta mahdollisesta geneettisen tiedon siirtymisestä proteiinista proteiiniin, RNA:han tai DNA:han pidetään mahdottomana. Tapaus prionien vaikutuksesta proteiineihin, jonka seurauksena samanlainen prioni muodostuu, voidaan ehdollisesti katsoa geneettisen informaation proteiinin → proteiinin toteutustyypeiksi. Muodollisesti se ei kuitenkaan ole sellainen, koska se ei vaikuta proteiinin aminohapposekvenssiin.
Käsitteen "keskusdogma" syntyhistoria on utelias. Koska sana dogma tarkoittaa yleensä lausuntoa, jota ei voi epäillä, ja sanalla itsessään on selkeä uskonnollinen konnotaatio, valitse se kuvaukseksi tieteellinen tosiasia ei täysin laillista. Francis Crickin itsensä mukaan se oli hänen virheensä. Hän halusi antaa esitetylle teorialle enemmän merkitystä, erottaa sen muiden teorioiden ja hypoteesien taustasta; miksi hän päätti käyttää tätä majesteettista, hänen mielestään sanaa ymmärtämättä sen todellista merkitystä. Nimi jäi kuitenkin kiinni.
Molekyylibiologia tänään
Molekyylibiologian nopea kehitys, yhteiskunnan jatkuva kiinnostus alan saavutuksiin ja tutkimuksen objektiivinen merkitys ovat johtaneet syntymiseen. suuri numero tärkeimmät molekyylibiologian tutkimuskeskukset ympäri maailmaa. Suurimmista mainittakoon seuraavat: molekyylibiologian laboratorio Cambridgessa, Royal Institute Lontoossa - Iso-Britanniassa; molekyylibiologian instituutit Pariisissa, Marseillessa ja Strasbourgissa, Pasteur-instituutti - Ranskassa; molekyylibiologian laitokset Harvardin yliopistossa ja Massachusetts Institute of Technologyssa, Berkeleyn yliopistossa, Kalifornian teknologiainstituutissa, Rockefeller-yliopistossa, Bethesdan kansanterveysinstituutissa - Yhdysvalloissa; Max Planck -instituutit, Göttingenin ja Münchenin yliopistot, Berliinin molekyylibiologian keskusinstituutti, Jenan ja Hallen instituutit - Saksassa; Karolinska-instituutti Tukholmassa, Ruotsissa.
Venäjällä tämän alan johtavat keskukset ovat molekyylibiologian instituutti. Institute of Molecular Genetics RAS, Institute of Gene Biology RAS, Institute of Physicochemical Biology nimetty V.A. A. N. Belozersky Moskovan valtionyliopisto. M.V. Lomonosov Biokemian instituutti. A.N. Bach RAS ja Protein RAS -instituutti Pushchinossa.
Nykyään molekyylibiologien kiinnostusalue kattaa laajan kirjon tieteellisiä peruskysymyksiä. Kuten ennenkin, johtavassa roolissa on nukleiinihappojen rakenteen ja proteiinien biosynteesin tutkiminen, erilaisten solunsisäisten rakenteiden ja solupintojen rakenteen ja toimintojen tutkiminen. Tärkeitä tutkimusalueita ovat myös vastaanotto- ja signaalinsiirtomekanismien, yhdisteiden kuljetuksen molekyylimekanismit solun sisällä ja myös solusta ulkoiseen ympäristöön ja takaisin. Sovellettavan molekyylibiologian tieteellisen tutkimuksen pääsuuntauksista yksi tärkeimmistä on kasvainten syntymisen ja kehityksen ongelma. Myös erittäin tärkeä alue, jota molekyylibiologian - molekyyligenetiikan -osasto tutkii, on esiintymisen molekyyliperustojen tutkiminen. perinnölliset sairaudet ja virustaudit, kuten AIDS, sekä menetelmien kehittäminen niiden ehkäisyyn ja mahdollisesti geenitasolla hoitoon. Molekyylibiologien löydöt ja kehitys oikeuslääketieteessä ovat löytäneet laajan sovelluksen. Venäjän, Yhdysvaltojen ja Iso-Britannian tutkijat tekivät 80-luvulla todellisen vallankumouksen henkilötunnistuksen alalla "genomisen sormenjälkien" - DNA:n tunnistamisen jokapäiväisessä käytännössä - menetelmän kehittämisen ja käyttöönoton ansiosta. Tämän alan tutkimus ei pysähdy tähän päivään asti, nykyaikaisten menetelmien avulla voidaan määrittää henkilö, jonka virhetodennäköisyys on prosentin miljardisosa. Geneettisen passin hanketta kehitetään jo aktiivisesti, mikä odotetusti vähentää rikollisuuden määrää huomattavasti.
Metodologia
Nykyään molekyylibiologialla on laaja arsenaali menetelmiä, joilla ratkaistaan edistyneimmät ja monimutkaisimmat tutkijoiden kohtaamat ongelmat.
Yksi yleisimmistä menetelmistä molekyylibiologiassa on geelielektroforeesi, joka ratkaisee ongelman makromolekyylien seoksen erottamisesta koon tai varauksen mukaan. Lähes aina geelissä olevien makromolekyylien erottamisen jälkeen käytetään blottausta, menetelmää, jonka avulla voit siirtää makromolekyylejä geelistä (sorb) kalvon pinnalle niiden kanssa tehtävän jatkotyöskentelyn, erityisesti hybridisaation, helpottamiseksi. Hybridisaatio - hybridi-DNA:n muodostaminen kahdesta eri luonteisesta säikeestä - menetelmä, jolla on tärkeä rooli perustutkimuksessa. Sitä käytetään määrittämään täydentäviä segmenttejä eri DNA:ssa (eri lajien DNA), sitä käytetään uusien geenien etsimiseen, sen avulla löydettiin RNA-interferenssi ja sen periaate muodosti genomisen sormenjälkien oton perustan.
Tärkeä rooli molekyylibiologisen tutkimuksen nykyaikaisessa käytännössä on sekvensointimenetelmällä - nukleiinihappojen nukleotidien ja proteiinien aminohappojen sekvenssin määrittämisellä.
Nykyaikaista molekyylibiologiaa ei voida kuvitella ilman polymeraasiketjureaktiomenetelmää (PCR). Tämän menetelmän ansiosta tietyn DNA-sekvenssin kopioiden lukumäärää (monistusta) lisätään, jotta yhdestä molekyylistä saadaan riittävä määrä ainetta jatkotyöskentelyyn sen kanssa. Samanlainen tulos saavutetaan molekyylikloonaustekniikalla, jossa vaadittu nukleotidisekvenssi viedään bakteerien (elävien järjestelmien) DNA:han, minkä jälkeen bakteerien lisääntyminen johtaa haluttuun tulokseen. Tämä lähestymistapa on teknisesti paljon monimutkaisempi, mutta sen avulla voidaan samanaikaisesti saada tulos tutkitun nukleotidisekvenssin ilmentymisestä.
Myös ultrasentrifugointimenetelmiä (makromolekyylien (suuret määrät), solujen, organellien erottamiseen), elektroni- ja fluoresenssimikroskopiaa, spektrofotometrisiä menetelmiä, röntgendiffraktioanalyysiä, autoradiografiaa jne. käytetään laajalti molekyylibiologisissa tutkimuksissa.
Teknologisen kehityksen ja kemian, fysiikan, biologian ja tietojenkäsittelytieteen alan tieteellisen tutkimuksen ansiosta nykyaikaiset laitteet mahdollistavat yksittäisten geenien ja niihin liittyvien prosessien eristämisen, tutkimisen ja muuttamisen.
1. Esittely.
Molekyylibiologian ja genetiikan oppiaine, tehtävät ja menetelmät. "Klassisen" genetiikan ja mikro-organismien genetiikan merkitys molekyylibiologian ja geenitekniikan kehityksessä. Geenin käsite "klassisessa" ja molekyyligenetiikassa, sen evoluutio. Geenitekniikan metodologian panos molekyyligenetiikan kehitykseen. Geenitekniikan soveltava arvo bioteknologiassa.
2. Perinnöllisyyden molekyyliperustat.
Solun käsite, sen makromolekyylikoostumus. Geneettisen materiaalin luonne. Historia todisteita DNA:n geneettisestä toiminnasta.
2.1. Erilaisia nukleiinihappoja. biologiset toiminnot nukleiinihapot. Kemiallinen rakenne, tilarakenne ja fyysiset ominaisuudet nukleiinihapot. Pro- ja eukaryoottien geneettisen materiaalin rakenteelliset ominaisuudet. Täydentävät Watson-Crick-emäsparit. Geneettinen koodi. Geneettisen koodin purkamisen historia. Koodin tärkeimmät ominaisuudet: tripletti, koodi ilman pilkkuja, degeneraatio. Koodisanakirjan ominaisuudet, kodoniperheet, semanttiset ja "merkittämättömät" kodonit. Pyöreät DNA-molekyylit ja DNA:n superkiertymisen käsite. DNA:n topoisomeerit ja niiden tyypit. Topoisomeraasien toimintamekanismit. Bakteerien DNA-gyraasi.
2.2. DNA:n transkriptio. Prokaryoottinen RNA-polymeraasi, sen alayksikkö ja kolmiulotteiset rakenteet. Erilaisia sigmatekijöitä. Prokaryoottigeenin promoottori rakenneosat. Transkriptiosyklin vaiheet. Transkription aloitus, "avoimen kompleksin" muodostuminen, venyminen ja lopettaminen. transkription vaimennus. Tryptofaanioperoniekspression säätely. "Riboswitches". Transkription lopetusmekanismit. Transkription negatiivinen ja positiivinen säätely. laktoosi operoni. Transkription säätely lambda-faagikehityksessä. DNA:n tunnistamisen periaatteet säätelyproteiinien (CAP-proteiini ja lambda-faagirepressori) avulla. Eukaryoottien transkription ominaisuudet. RNA:n prosessointi eukaryooteissa. Transkriptien päättäminen, silmukointi ja polyadenylaatio. liitosmekanismit. Pienen tuman RNA:n ja proteiinitekijöiden rooli. Vaihtoehtoinen liitos, esimerkkejä.
2.3. Lähettää, sen vaiheet, ribosomien toiminta. Ribosomien sijainti solussa. Prokaryoottiset ja eukaryoottiset ribosomityypit; 70S ja 80S ribosomit. Ribosomien morfologia. Jako alahiukkasiin (alayksiköihin). Kodoniriippuvainen aminoasyyli-tRNA:n sitoutuminen elongaatiosyklissä. Kodoni-antikodoni vuorovaikutus. Elongaatiotekijän EF1 (EF-Tu) osallistuminen aminoasyyli-tRNA:n sitoutumiseen ribosomiin. Venymätekijä EF1B (EF-Ts), sen toiminta, reaktiosarja sen osallistumisen kanssa. Antibiootit, jotka vaikuttavat aminoasyyli-tRNA:n kodonista riippuvaisen sitoutumisen vaiheeseen ribosomiin. Aminoglykosidiantibiootit (streptomysiini, neomysiini, kanamysiini, gentamysiini jne.), niiden vaikutusmekanismi. Tetrasykliinit aminoasyyli-tRNA:n ribosomiin sitoutumisen estäjinä. Lähetyksen aloitus. Aloitusprosessin päävaiheet. Translaation aloitus prokaryooteissa: aloitustekijät, aloituskodonit, pienen ribosomaalisen alayksikön RNA:n 3¢-pää ja Shine-Dalgarno-sekvenssi mRNA:ssa. Translaation aloitus eukaryooteissa: aloitustekijät, aloituskodonit, 5¢-transloitumaton alue ja cap-riippuvainen terminaalinen aloitus. "Sisäinen" cap-riippumaton aloitus eukaryooteissa. Transpeptidaatio. Transpeptidaation estäjät: kloramfenikoli, linkomysiini, amisetiini, streptogramiinit, anisomysiini. Translokaatio. Venymäkertoimen EF2 (EF-G) ja GTP:n osallisuus. Translokaatioestäjät: fusidiinihappo, viomysiini, niiden vaikutusmekanismit. Käännöksen lopettaminen. Lopetuskodonit. Prokaryoottien ja eukaryoottien proteiinin lopetustekijät; kaksi lopetustekijöiden luokkaa ja niiden toimintamekanismit. Käännöksen säätely prokaryooteissa.
2.4. DNA kopiointi ja sen geneettinen hallinta. Replikaatioon osallistuvat polymeraasit, niiden entsymaattisten toimintojen ominaisuudet. DNA:n uskollisuus. DNA-emäsparien välisten steeristen vuorovaikutusten rooli replikaation aikana. E. coli -polymeraasit I, II ja III. Polymeraasi III -alayksiköt. Replikointihaarukka, "johtavat" ja "jäävät" säikeet replikoinnin aikana. Fragmentteja Okazakista. Proteiinikompleksi replikaatiohaarukassa. Replikaation aloituksen säätely E. colissa. Replikaation lopettaminen bakteereissa. Plasmidin replikaation säätelyn piirteet. Kaksisuuntainen ja pyörivä renkaan replikointi.
2.5. Rekombinaatio, sen tyypit ja mallit. Yleinen tai homologinen rekombinaatio. Kaksijuosteiset katkokset DNA:ssa, jotka käynnistävät rekombinaation. Rekombinaation rooli kaksisäikeisten katkeamien replikaation jälkeisessä korjauksessa. Holliday-rakenne rekombinaatiomallissa. Yleisen rekombinaation entsymologia E. colissa. RecBCD-kompleksi. Reca-proteiini. Rekombinaation rooli DNA-synteesin varmistamisessa replikaation keskeyttävissä DNA-vaurioissa. rekombinaatio eukaryooteissa. Rekombinaatioentsyymit eukaryooteissa. Paikkakohtainen rekombinaatio. Erot yleisen ja paikkaspesifisen rekombinaation molekyylimekanismeissa. Rekombinaasien luokitus. Paikkaspesifisen rekombinaation aikana suoritettujen kromosomien uudelleenjärjestelyjen tyypit. Paikkaspesifisen rekombinaation säätelyrooli bakteereissa. Monisoluisten eukaryoottisten kromosomien rakentaminen käyttämällä paikkaspesifistä faagirekombinaatiojärjestelmää.
2.6. DNA:n korjaus. Korjaustyyppien luokittelu. Tymiinidimeerien ja metyloidun guaniinin suora korjaus. Pohjien leikkaaminen. Glykosylaasit. Pariutumattomien nukleotidien korjausmekanismi (epäsopivuuskorjaus). Korjattavan DNA-juosteen valinta. SOS korjaus. Prokaryoottien ja eukaryoottien SOS-korjaukseen osallistuvien DNA-polymeraasien ominaisuudet. Käsite " adaptiivisia mutaatioita”bakteereissa. Kaksijuosteisten katkeamien korjaaminen: homologinen replikatiivinen rekombinaatio ja DNA-molekyylin ei-homologisten päiden yhdistäminen. Replikaatio-, rekombinaatio- ja korjausprosessien välinen suhde.
3. Mutaatioprosessi.
Biokemiallisten mutanttien rooli yhden geenin - yhden entsyymin teorian muodostumisessa. Mutaatioluokitus. Pistemutaatiot ja kromosomien uudelleenjärjestelyt, niiden muodostumismekanismi. Spontaani ja indusoitu mutageneesi. Mutageenien luokitus. Mutageneesin molekyylimekanismi. Mutageneesin ja korjauksen välinen suhde. Mutanttien tunnistaminen ja valinta. Suppressio: intrageeninen, intergeeninen ja fenotyyppinen.
4. Ekstrakromosomaalinen geneettisiä elementtejä.
Plasmidit, niiden rakenne ja luokittelu. Sukupuolitekijä F, sen rakenne ja elinkaari. Tekijän F rooli kromosominsiirron mobilisaatiossa. Hfr- ja F-luovuttajien muodostuminen Konjugaatiomekanismi Bakteriofagit, niiden rakenne ja elinkaari Virulentit ja lauhkeat bakteriofagit Lysogenia ja transduktio Yleinen ja spesifinen transduktio Migroituvat geneettiset elementit: transposonit ja IS-sekvenssit, niiden rooli geneettisessä aineenvaihdunnassa DNA - transposonit prokaryoottien ja eukaryoottien genomissa Bakteerien IS-sekvenssit, niiden rakenne IS-sekvenssit bakteerien F-tekijän komponenttina, joka määrää kyvyn siirtää geneettistä materiaalia konjugoinnin aikana Bakteerien ja eukaryoottisten organismien transposonit Suora replikoitumaton ja transpositioiden replikatiiviset mekanismit Horisontaalisen transposonisiirron käsite ja niiden rooli rakenteellisissa uudelleenjärjestelyissä (ektooppinen rekombinaatio) ja genomin evoluutiossa.
5. Geenin rakenteen ja toiminnan tutkimus.
Geneettisen analyysin elementit. Cis-trans-komplementaatiotesti. Geneettinen kartoitus konjugaatiolla, transduktiolla ja transformaatiolla. Geneettisten karttojen rakentaminen. Hieno geneettinen kartoitus. Geenirakenteen fyysinen analyysi. heterodupleksianalyysi. Rajoitusanalyysi. Sekvensointimenetelmät. polymeraasiketjureaktio. Geenin toiminnan paljastaminen.
6. Geeniekspression säätely. Käsitteet operonista ja regulonista. Kontrolli transkription aloitustasolla. Promoottori-, operaattori- ja säätelyproteiinit. Geeniekspression positiivinen ja negatiivinen kontrolli. Ohjaus transkription lopettamisen tasolla. Kataboliittikontrolloidut operonit: mallit laktoosi-, galaktoosi-, arabinoosi- ja maltoosioperoneista. Vaimentimella ohjatut operonit: tryptofaanioperonin malli. Geeniekspression moniarvoinen säätely. Globaalit sääntelyjärjestelmät. Sääntelyreaktio stressiin. transkription jälkeinen kontrolli. signaalin siirto. RNA-välitteinen säätely: pienet RNA:t, sensori-RNA:t.
7. Geenitekniikan perusteet. Restriktioentsyymit ja modifikaatiot. Geenien eristäminen ja kloonaus. Vektorit molekyylikloonausta varten. Yhdistelmä-DNA:n rakentamisen periaatteet ja niiden vieminen vastaanottajasoluihin. Geenitekniikan soveltavat näkökohdat.
A). Pääkirjallisuus:
1. Watson J., Tooze J., Recombinant DNA: A Brief Course. – M.: Mir, 1986.
2. Geenit. – M.: Mir. 1987.
3. Molekyylibiologia: nukleiinihappojen rakenne ja biosynteesi. /Toim. . - M. Korkeakoulu. 1990.
4., - Molekyylibiotekniikka. M. 2002.
5. Spiriiniribosomit ja proteiinien biosynteesi. – M.: valmistua koulusta, 1986.
b). Lisäkirjallisuutta:
1. Genomin hesiini. – M.: Tiede. 1984.
2. Geenitekniikan Rybchin. - Pietari: Pietarin valtion teknillinen yliopisto. 1999.
3. Patruševin geenit. – M.: Nauka, 2000.
4. Moderni mikrobiologia. Prokaryootit (2 osassa). – M.: Mir, 2005.
5. M. Singer, P. Berg. Geenit ja genomit. – M.: Mir, 1998.
6. Shchelkunov-tekniikka. - Novosibirsk: Sibistä. Yliopisto, 2004.
7. Stepanov-biologia. Proteiinien rakenne ja toiminta. - M.: V. Sh., 1996.
haastatella
Pirogov Sergey - osallistuja "Norsun ja kirahvin" järjestämän biologian olympialaisten valmisteluun vuonna 2012.
Kansainvälisen biologian Universiadin voittaja
Olympian "Lomonosov" voittaja
Alueellisen vaiheen voittaja Koko Venäjän olympialaiset biologiassa vuonna 2012
Opiskelu Moskovan valtionyliopistossa. M.V. Lomonosov Biologian tiedekunnassa: Molekyylibiologian laitos, 6. vuoden opiskelija. Työskentelee molekyyligenetiikan instituutin eläinten biokemiallisen genetiikan laboratoriossa.
- Seryozha, jos lukijoilla on kysyttävää, voivatko he kysyä sinulta?
Kyllä, voit tietysti esittää kysymyksiä ainakin välittömästi. Tällä alalla:
Napsauta tätä esittääksesi kysymyksen.

- Aloitetaan koulusta, eikö sinulla ollut superhieno koulu?
Opiskelin erittäin heikolla Moskovan koulussa, sellaisessa keskivertokoulussa. Totta, meillä oli upea opettaja Moskovan taideteatterissa, jonka ansiosta meillä oli suurelta osin nimellinen koulun "taidehistoriallinen" suunta.
- Entä biologia?
Biologian opettajamme oli hyvin iäkäs, kuuro ja terävä nainen, jota kaikki pelkäsivät. Mutta rakkaus aiheeseensa ei lisännyt. Olen ollut intohimoisesti biologiasta lapsesta asti, viisivuotiaasta lähtien. Luen itse kaiken, lähinnä anatomia ja eläintiede. Niin koulutarvikkeet olemassa rinnakkain omien etujeni kanssa. Olympialaiset muuttivat kaiken.
- Kerro minulle lisää siitä.
7. luokalla osallistuin ensimmäistä kertaa kunnalliseen vaiheeseen (tietysti melkein kaikissa aineissa kerralla, koska olin ainoa oppilas, jonka opettajilla oli syytä lähettää). Ja hän voitti biologiassa. Sitten koulu piti tätä hauskana, mutta ei kovin mielenkiintoisena tosiasiana.

- Auttoiko se sinua koulussa?
Muistan, että loistavista opiskeluistani huolimatta sain usein biologian opettajalta B:n näppylällä kuten "sipulin osan piirustuksessa juuret tulee maalata ruskeiksi, ei harmaiksi." Kaikki oli aika masentavaa. Kahdeksannella luokalla menin jälleen olympialaisiin, mutta jostain syystä minua ei lähetetty biologiaan. Mutta hänestä tuli voittaja ja palkinnon voittaja muissa aiheissa.
- Mitä tapahtui 9. luokalla?
9. luokalla en mennyt piirivaiheeseen. Siellä sain yllättäen heikon, raja-arvosanan, joka kuitenkin osoittautui aluevaiheelle siirtymiseksi. Sillä oli voimakas motivoiva voima - ymmärrys siitä, kuinka paljon en tiedä ja kuinka moni tietää kaiken tämän (kuinka monta tällaista ihmistä valtakunnallisessa mittakaavassa pelkäsin edes kuvitella).

- Kerro meille, kuinka valmistauduit.
Intensiivinen itseopiskelu, kirjakauppojen tutkailut ja tuhannet viime vuoden tehtävät vaikuttivat parantavasti. Sain yhden teorian korkeimmista pisteistä (joka oli myös minulle täysin odottamaton), menin käytännön vaiheeseen ... ja epäonnistuin. Tuolloin en edes tiennyt käytännön vaiheen olemassaolosta.
- Vaikuttiko olympialaiset sinuun?
Elämäni on muuttunut radikaalisti. Opin monista muista olympialaisista, erityisesti rakastuin SBO:hen. Myöhemmin hän näytti monia hyvät tulokset, jotkut voittivat, kiitos "Lomonosovskaya" sai oikeuden tulla ilman kokeita. Samaan aikaan voitin taidehistorian olympialaisia, joihin hengitän edelleen epätasaisesti. Totta, hän ei ollut ystävä käytännön matkojen kanssa. 11. luokalla pääsin kuitenkin loppuvaiheeseen, mutta Fortune ei ollut suotuisa, enkä tällä kertaa ehtinyt täyttää teoreettisen vaiheen vastausmatriisia. Mutta tämä mahdollisti sen, että käytännöllisyydestä ei tarvinnut huolehtia liikaa.
- Oletko tavannut monia olympialaisia?
Kyllä, olen edelleen sitä mieltä, että olin erittäin onnekas ikätovereideni kanssa, jotka laajensivat suuresti näköalojani. Olympialaisten toinen puoli, motivaation lisäksi opiskella aihetta harmonisemmin, oli tutustuminen olympialaisiin. Huomasin jo tuolloin, että horisontaalinen kommunikaatio on joskus hyödyllisempää kuin vertikaalinen kommunikointi - opettajien kanssa harjoitusleirillä.

- Miten pääsit yliopistoon? Valitsitko tiedekunnan?
11. luokan jälkeen astuin Moskovan valtionyliopiston biologian tiedekuntaan. Vain suurin osa silloisista tovereistani teki valinnan FBB:n hyväksi, mutta tässä päärooli oli sillä, että minusta ei tullut All-Russianin voittajaa. Joten minun piti tehdä sisäinen koe matematiikasta, ja siinä, varsinkin koulussa - rakastuin ylempään paljon enemmän - en ollut vahva. Ja koulussa oli erittäin huono valmistautuminen (emme olleet edes valmistautuneet melkein koko C-osaan). Kiinnostuksen suhteen arvelin jo silloin, että lopulta voit päästä mihin tahansa tulokseen sisäänpääsypaikasta riippumatta. Myöhemmin kävi ilmi, että monet FBB:stä valmistuneet ovat siirtyneet pääosin märkäbiologiaan ja päinvastoin - monet hyvät bioinformaatikot aloittivat amatöörinä. Vaikka sillä hetkellä minusta tuntui, että biologisen tiedekunnan osasto olisi erilainen kuin FBBshny. Tässä olin varmasti väärässä.
Tiesitkö?
Mielenkiintoista
Tiesitkö?
Mielenkiintoista
Elefantti ja kirahvi -leirillä on vuoroja biokemiassa ja molekyylibiologiassa, jossa koululaiset yhdessä Moskovan valtionyliopiston kokeneiden opettajien kanssa perustavat kokeita ja valmistautuvat olympialaisiin.© Haastatteli Reshetov Denis. Valokuvat tarjosi ystävällisesti Sergei Pirogov.
Nukleiinihappojen ja proteiinien biosynteesin tutkimuksen edistyminen on johtanut useiden menetelmien luomiseen, joilla on suuri käytännön merkitys lääketieteessä, maataloudessa ja useilla muilla teollisuudenaloilla.
Geneettisen koodin sekä perinnöllisen tiedon tallennuksen ja toteuttamisen perusperiaatteiden tutkimisen jälkeen molekyylibiologian kehitys pysähtyi, koska ei ollut menetelmiä, jotka olisivat mahdollistaneet geenien manipulointia, eristämistä ja muuttamista. Nämä menetelmät syntyivät 1970-1980-luvuilla. Tämä antoi voimakkaan sysäyksen tämän edelleen kukoistavan tieteenalan kehitykselle. Ensinnäkin nämä menetelmät koskevat yksittäisten geenien hankkimista ja niiden viemistä muiden organismien soluihin (molekyylikloonaus ja siirtogeneesi, PCR) sekä menetelmiä geenien nukleotidisekvenssin määrittämiseksi (DNA- ja RNA-sekvensointi). Näitä menetelmiä käsitellään yksityiskohtaisemmin alla. Aloitamme yksinkertaisimmalla perusmenetelmällä, elektroforeesilla, ja siirrymme sitten monimutkaisempiin menetelmiin.
DNA ELEKTROFOREESI
Se on DNA:n kanssa työskentelyn perusmenetelmä, jota käytetään lähes kaikkien muiden menetelmien ohella haluttujen molekyylien eristämiseen ja tulosten analysointiin. Geelielektroforeesia käytetään DNA-fragmenttien erottamiseen pituuden mukaan. DNA on happo, sen molekyylit sisältävät fosforihappojäämiä, jotka irrottavat protonin ja saavat negatiivisen varauksen (kuva 1).
Siksi sähkökentässä DNA-molekyylit liikkuvat kohti anodia - positiivisesti varautunutta elektrodia. Tämä tapahtuu varauksenkantaja-ioneja sisältävässä elektrolyyttiliuoksessa, minkä vuoksi tämä liuos johtaa virtaa. Fragmenttien erottamiseen käytetään tiheää polymeereistä (agaroosista tai polyakryyliamidista) valmistettua geeliä. DNA-molekyylit "kietoutuvat" siihen mitä enemmän, sitä pidempiä ne ovat, ja siksi pisimmät molekyylit liikkuvat hitaimmin ja lyhyimmät - nopeimmin (kuva 2). Ennen elektroforeesia tai sen jälkeen geeliä käsitellään väriaineilla, jotka sitoutuvat DNA:han ja fluoresoivat ultraviolettivalossa, ja geelissä saadaan vyöhykekuvio (katso kuva 3). Näytteessä olevien DNA-fragmenttien pituuksien määrittämiseksi niitä verrataan markkeriin, ts. sarjaan standardipituisia fragmentteja, jotka on kerrostettu rinnakkain samalle geelille (kuvio 4).



Tärkeimmät työkalut DNA:n kanssa työskentelyyn ovat entsyymit, jotka suorittavat DNA-transformaatioita elävissä soluissa: DNA-polymeraasit, DNA-ligaasit ja restriktioendonukleaasit eli restriktioentsyymit. DNA-polymeraasi Suoritetaan DNA-templaattisynteesi, joka mahdollistaa DNA:n lisäämisen koeputkessa. DNA-ligaasit ompele DNA-molekyylejä yhteen tai korjaa niissä olevat aukot. Restriktioendonukleaasit, tai rajoittaa, leikkaa DNA-molekyylejä tiukasti määriteltyjen sekvenssien mukaan, mikä mahdollistaa yksittäisten fragmenttien leikkaamisen DNA:n kokonaismassasta. Nämä fragmentit voivat joissakin tapauksissa sisältää yksittäisiä geenejä.
rajoittaa
Restriktioentsyymien tunnistamat sekvenssit ovat symmetrisiä, ja katkoksia voi tapahtua sellaisen sekvenssin keskellä tai siirtymällä (samassa paikassa molemmissa DNA-juosteissa). Toimintasuunnitelma eri tyyppejä restrikaasi on esitetty kuvassa. 1. Ensimmäisessä tapauksessa saadaan niin sanotut "tylpät" päät ja toisessa - "tahmeat" päät. Pohjan "tahmeiden" päiden tapauksessa ketju on toista lyhyempi, muodostuu yksisäikeinen osa symmetrisellä sekvenssillä, joka on sama molemmissa muodostetuissa päissä.
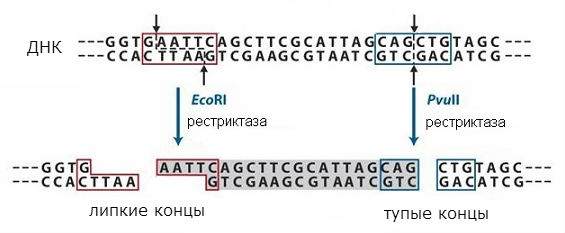
Päätesekvenssit ovat samat, kun mikä tahansa DNA katkaistaan tietyllä restriktioentsyymillä, ja ne voidaan yhdistää uudelleen, koska niillä on komplementaarisia sekvenssejä. Ne voidaan liittää DNA-ligaasin kanssa yhden molekyylin muodostamiseksi. Siten on mahdollista yhdistää kahden eri DNA:n fragmentteja ja saada ns yhdistelmä-DNA. Tätä lähestymistapaa käytetään molekyylikloonausmenetelmässä, joka mahdollistaa yksittäisten geenien saamisen ja viemisen soluihin, jotka voivat muodostaa geenin koodaaman proteiinin.
molekyylikloonaus
Molekyylikloonaus käyttää kahta DNA-molekyyliä - inserttiä, joka sisältää kiinnostavan geenin, ja vektori- DNA toimii kantajana. Insertti "ommellaan" vektoriin entsyymien avulla, jolloin saadaan uusi, yhdistelmä-DNA-molekyyli, sitten tämä molekyyli viedään isäntäsoluihin, ja nämä solut muodostavat pesäkkeitä ravintoalustalle. Pesäke on yhden solun jälkeläinen eli klooni, kaikki pesäkkeen solut ovat geneettisesti identtisiä ja sisältävät saman rekombinantti-DNA:n. Tästä johtuu termi "molekyylikloonaus", toisin sanoen meitä kiinnostavan DNA-fragmentin sisältävän solukloonin hankkiminen. Kun meitä kiinnostavan insertin sisältävät pesäkkeet on saatu, on mahdollista karakterisoida tämä insertti eri menetelmillä, esimerkiksi määrittää sen tarkka sekvenssi. Solut voivat myös tuottaa insertin koodaamaa proteiinia, jos se sisältää toiminnallisen geenin.
Kun rekombinanttimolekyyli viedään soluihin, tapahtuu näiden solujen geneettinen transformaatio. Muutos- prosessi, jossa organismin solu absorptioi ympäristöstä vapaan DNA-molekyylin ja integroituu genomiin, mikä johtaa siihen, että tällaisessa solussa ilmaantuu uusia periytyviä piirteitä, jotka ovat ominaisia DNA:n luovuttajalle. . Esimerkiksi, jos lisätty molekyyli sisältää geenin, joka on vastustuskykyinen antibiootille ampisilliinille, transformoidut bakteerit kasvavat sen läsnä ollessa. Ennen transformaatiota ampisilliini aiheutti heidän kuolemansa, toisin sanoen transformoituneisiin soluihin ilmestyy uusi merkki.
VEKTORIT
Vektorilla on oltava useita ominaisuuksia:
Ensinnäkin se on suhteellisen pieni DNA-molekyyli, jota on helppo manipuloida.
Toiseksi, jotta DNA säilyisi ja lisääntyisi solussa, sen täytyy sisältää tietty sekvenssi, joka varmistaa sen replikaation (replikaation aloituskohta tai replikaation aloituskohta).
Kolmanneksi sen on sisällettävä markkerigeeni, joka varmistaa vain niiden solujen valinnan, joihin vektori on tullut. Yleensä nämä ovat antibioottiresistenssigeenejä - sitten antibiootin läsnä ollessa kaikki solut, jotka eivät sisällä vektoria, kuolevat.
Geenikloonaus suoritetaan useimmiten bakteerisoluissa, koska niitä on helppo viljellä ja ne lisääntyvät nopeasti. Bakteerisolussa on yleensä yksi suuri pyöreä, useita miljoonia emäspareja pitkä DNA-molekyyli, joka sisältää kaikki bakteereille tarpeelliset geenit - bakteerikromosomi. Sen lisäksi joissakin bakteereissa on pieniä (useita tuhatta emäsparia) pyöreää DNA:ta, ns plasmidit(Kuva 2). Ne, kuten pää-DNA, sisältävät nukleotidisekvenssin, joka tarjoaa DNA:lle kyvyn replikoitua (ori). Plasmidit replikoituvat pääasiallisesta (kromosomaalisesta) DNA:sta riippumatta, joten ne ovat läsnä solussa suurissa määrissä kopioita. Monet näistä plasmideista sisältävät antibioottiresistenssigeenejä, mikä mahdollistaa plasmidia kantavien solujen erottamisen normaaleista soluista. Yleisemmin käytetään plasmideja, joissa on kaksi geeniä, jotka antavat resistenssin kahdelle antibiootille, kuten tetrasykliinille ja amysiliinille. Olla olemassa yksinkertaisia menetelmiä sellaisen plasmidi-DNA:n eristäminen, joka on vapaa bakteerin pääkromosomin DNA:sta.

TRANSGENEESIIN MERKITYS
Geenien siirtymistä organismista toiseen kutsutaan transgeneesi ja sellaiset muunnetut organismit - siirtogeeninen. Geeninsiirtomenetelmää mikrobisoluihin käytetään yhdistelmä-DNA-tekniikalla valmistettujen proteiinivalmisteiden saamiseksi lääketieteelliseen käyttöön, erityisesti ihmisen proteiineja, jotka eivät aiheuta immuunihyljintää - interferoneja, insuliinia ja muita proteiinihormoneja, solujen kasvutekijöitä sekä proteiineja rokotteet. Monimutkaisemmissa tapauksissa, kun proteiinin modifiointi suoritetaan oikein vain eukaryoottisoluissa, käytetään siirtogeenisiä soluviljelmiä tai siirtogeenisiä eläimiä, erityisesti karjaa (pääasiassa vuohia), jotka erittävät tarvittavat proteiinit maitoon tai eristetään proteiineja niiden verestä. . Näin saadaan vasta-aineita, veren hyytymistekijöitä ja muita proteiineja. saatu transgeneesillä viljellyt kasvit jotka kestävät rikkakasvien torjunta-aineita ja tuholaisia ja joilla on muita hyödyllisiä ominaisuuksia. Siirtogeenisiä mikro-organismeja käyttämällä jäteveden puhdistamiseen ja saastumisen torjuntaan on jopa siirtogeenisiä mikrobeja, jotka voivat hajottaa öljyä. Lisäksi siirtogeeniset teknologiat ovat välttämättömiä tieteellinen tutkimus- Biologian kehitystä nykyään ei voida ajatella ilman muunnos- ja geeninsiirtomenetelmien rutiininomaista käyttöä.
molekyylikloonaustekniikka
insertit
Yksittäisen geenin saamiseksi mistä tahansa organismista kaikki kromosomaalinen DNA eristetään siitä ja pilkotaan yhdellä tai kahdella restriktioentsyymillä. Entsyymit valitaan siten, että ne eivät leikkaa meitä kiinnostavaa geeniä, vaan tekevät katkoja sen reunoille ja plasmidi-DNA:ssa yhden katkaisun johonkin resistenssigeeniin esimerkiksi ampisilliinille.
Molekyylikloonausprosessi sisältää seuraavat vaiheet:
Leikkaa ja ompele - yhden rekombinanttimolekyylin rakentaminen insertistä ja vektorista.
Transformaatio on rekombinanttimolekyylin viemistä soluihin.
Valinta - solujen valinta, jotka ovat vastaanottaneet vektorin, jossa on insertti.
leikkaus ja ompeleminen
Plasmidi-DNA:ta käsitellään samoilla restriktioentsyymeillä ja se muuttuu lineaariseksi molekyyliksi, jos valitaan sellainen restriktioentsyymi, joka tuo plasmidiin yhden katkoksen. Tämän seurauksena samat tahmeat päät näkyvät kaikkien tuloksena olevien DNA-fragmenttien päissä. Kun lämpötilaa lasketaan, nämä päät yhdistyvät satunnaisesti ja ligoidaan DNA-ligaasilla (katso kuvio 3).

Saadaan seos koostumukseltaan erilaisia pyöreitä DNA:ita: osa niistä sisältää tietyn DNA-sekvenssin kromosomaalista DNA:ta yhdistettynä bakteeri-DNA:han, toiset sisältävät kromosomaalisen DNA:n fragmentteja toisiinsa liittyneenä ja toiset sisältävät pelkistetyn pyöreän plasmidin tai sen dimeerin. (Kuva 4).

muunnos
Seuraavaksi tämä seos suoritetaan geneettinen transformaatio bakteerit, jotka eivät sisällä plasmideja. Muutos- prosessi, jossa organismin solu absorptioi ympäristöstä vapaan DNA-molekyylin ja integroituu genomiin, mikä johtaa siihen, että tällaisessa solussa ilmaantuu uusia periytyviä piirteitä, jotka ovat ominaisia DNA:n luovuttajalle. . Vain yksi plasmidi voi päästä sisään ja lisääntyä jokaisessa solussa. Tällaiset solut asetetaan kiinteälle ravintoalustalle, joka sisältää antibiootti tetrasykliiniä. Solut, jotka eivät ole saaneet plasmidia, eivät kasva tällä alustalla, ja plasmidia kantavat solut muodostavat pesäkkeitä, joista jokainen sisältää vain yhden solun jälkeläisiä, ts. kaikki solut pesäkkeessä kantavat samaa plasmidia (katso kuvio 5).

Valinta
Seuraavaksi tehtävänä on eristää vain solut, joihin insertin sisältävä vektori on mennyt, ja erottaa ne soluista, joissa on vain vektori ilman inserttiä tai jotka eivät sisällä vektoria ollenkaan. Tätä prosessia oikeiden solujen valitsemiseksi kutsutaan valinta. Tätä varten hae selektiiviset merkit- yleensä antibioottiresistenssigeenit vektorissa, ja valikoiva media jotka sisältävät antibiootteja tai muita selektiivisiä aineita.
Tarkastelemassamme esimerkissä ampisilliinin läsnä ollessa kasvatetuista pesäkkeistä peräisin olevia soluja jatkoviljellään kahdella alustalla: ensimmäinen sisältää ampisilliinia ja toinen tetrasykliiniä. Pesäkkeet, jotka sisältävät vain plasmidin, kasvavat molemmilla elatusaineilla, kun taas pesäkkeet, jotka sisältävät insertoitua kromosomaalista DNA:ta plasmideissa, eivät kasva tetrasykliiniä sisältävällä alustalla (kuvio 5). Niistä ne, jotka sisältävät meitä kiinnostavan geenin, valitaan erityisillä menetelmillä, kasvatetaan riittävästi ja eristetään plasmidi-DNA. Siitä leikataan pois kiinnostava yksittäinen geeni käyttämällä samoja restriktaaseja, joita käytettiin rekombinantti-DNA:n saamiseksi. Tämän geenin DNA:ta voidaan käyttää nukleotidien sekvenssin määrittämiseen, viemiseen mihin tahansa organismiin uusien ominaisuuksien saamiseksi tai halutun proteiinin syntetisoimiseen. Tätä geenieristysmenetelmää kutsutaan molekyylikloonaus.
fluoresoivat proteiinit
On erittäin kätevää käyttää fluoresoivia proteiineja markkerigeeneinä eukaryoottisten organismien tutkimuksissa. Ensimmäisen fluoresoivan proteiinin geeni, vihreä fluoresoiva proteiini (GFP) eristettiin meduusa Aqeuorea victoriasta ja vietiin erilaisiin mallieliöihin (katso kuva 6) Nobel palkinto tämän proteiinin löytämiseen ja käyttöön.

Sitten eristettiin muiden fluoresoivien proteiinien - punaisen, sinisen, keltaisen - geenit. Näitä geenejä on modifioitu keinotekoisesti tuottamaan proteiineja, joilla on halutut ominaisuudet. Fluoresoivien proteiinien monimuotoisuus on esitetty kuvassa. 7, jossa on petrimalja, jossa on bakteereja, jotka sisältävät geenejä eri fluoresoiville proteiineille.

fluoresoivien proteiinien käyttö
Fluoresoiva proteiinigeeni voidaan fuusioida minkä tahansa muun proteiinin geeniin, jolloin translaation aikana muodostuu yksi proteiini - translaatiofuusioproteiini tai fuusio(fuusioproteiini), joka fluoresoi. Siten on mahdollista tutkia esimerkiksi minkä tahansa kiinnostavan proteiinin sijaintia (sijaintia) solussa, niiden liikkumista. Ilmentämällä fluoresoivia proteiineja vain tietyissä solutyypeissä on mahdollista leimata tämän tyyppisiä soluja monisoluinen organismi(katso kuva 8 - hiiren aivot, joissa yksittäisillä hermosoluilla on erilaiset värit tietyn fluoresoivien proteiinigeenien yhdistelmän vuoksi). Fluoresoivat proteiinit ovat välttämätön työkalu nykyaikaisessa molekyylibiologiassa.

PCR
Toista menetelmää geenien saamiseksi kutsutaan polymeraasiketjureaktio (PCR). Se perustuu DNA-polymeraasien kykyyn täydentää DNA:n toinen juoste komplementaarista juostetta pitkin, kuten tapahtuu soluissa DNA:n replikaation aikana.
Replikaation alkukohdat tässä menetelmässä saadaan kahdesta pienestä DNA-palasta, joita kutsutaan nimellä siemenet, tai alukkeet. Nämä alukkeet ovat komplementaarisia kiinnostuksen kohteena olevan geenin päille kahdessa DNA-juosteessa. Ensinnäkin kromosomaalinen DNA, josta geeni eristetään, sekoitetaan siementen kanssa ja kuumennetaan 99 °C:seen. Tämä johtaa vetysidosten katkeamiseen ja DNA-säikeiden erottumiseen. Sen jälkeen lämpötila lasketaan noin 50-70 asteeseen (riippuen siementen pituudesta ja järjestyksestä). Näissä olosuhteissa alukkeet kiinnittyvät kromosomaalisen DNA:n komplementaarisiin alueisiin muodostaen säännöllisen kaksoiskierteen (katso kuvio 9). Sen jälkeen lisätään kaikkien neljän DNA-synteesiin tarvittavan nukleotidin ja DNA-polymeraasin seos. Entsyymi pidentää alukkeita rakentamalla kaksijuosteista DNA:ta alukkeiden kiinnityskohdasta, ts. geenin päistä yksijuosteisen kromosomimolekyylin loppuun. 
Jos seosta kuumennetaan nyt uudelleen, kromosomaaliset ja vasta syntetisoidut ketjut hajaantuvat. Jäähtymisen jälkeen siemenet liittyvät taas niihin, joita otetaan runsaasti (katso kuva 10).

Äskettäin syntetisoiduissa ketjuissa ne eivät liity siihen päähän, josta ensimmäinen synteesi alkoi, vaan vastakkaiseen, koska DNA-ketjut ovat vastakkaisia. Siksi toisessa synteesisyklissä vain geeniä vastaava sekvenssi valmistuu tällaisissa ketjuissa (katso kuvio 11).
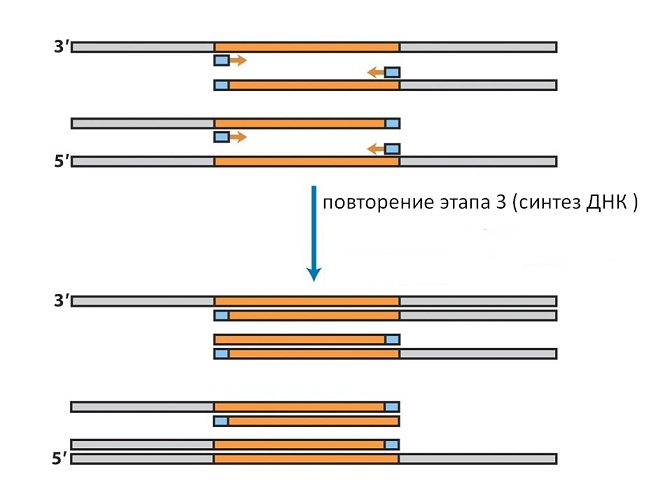
Tämä menetelmä käyttää DNA-polymeraasia termofiilisistä bakteereista, jotka kestävät kiehumista ja toimivat 70-80 °C:n lämpötiloissa, sitä ei tarvitse lisätä joka kerta, mutta riittää, kun se lisätään kokeen alussa. Toistamalla lämmitys- ja jäähdytystoimenpiteet samassa järjestyksessä, voimme kaksinkertaistaa jaksojen lukumäärän kussakin jaksossa, jota molemmista päistä rajoittavat lisätyt siemenet (katso kuva 12).

Noin 25 tällaisen syklin jälkeen geenin kopioiden määrä kasvaa yli miljoona kertaa. Tällaiset määrät voidaan helposti erottaa koeputkeen syötetystä kromosomaalisesta DNA:sta ja käyttää eri tarkoituksiin.
DNA-sekvensointi
Toinen tärkeä saavutus on menetelmien kehittäminen DNA:n nukleotidisekvenssin määrittämiseksi - DNA-sekvensointi(englanninkielisestä sekvenssistä - sekvenssi). Tätä varten on tarpeen saada geenit puhtaina toisesta DNA:sta jollakin kuvatuista menetelmistä. Sitten DNA-ketjut erotetaan kuumentamalla ja niihin lisätään radioaktiivisella fosforilla leimattu aluke tai fluoresoiva leima. Huomaa, että otetaan yksi siemen, joka täydentää yhtä ketjua. Sitten lisätään DNA-polymeraasi ja 4 nukleotidin seos. Tällainen seos jaetaan 4 osaan ja jokaiseen lisätään yksi nukleotideista modifioituna siten, että se ei sisällä hydroksyyliryhmää deoksiriboosin kolmannessa atomissa. Jos tällainen nukleotidi sisältyy syntetisoituun DNA-ketjuun, sen pidentyminen ei voi jatkua, koska polymeraasilla ei ole minnekään kiinnittää seuraavaa nukleotidia. Siksi DNA-synteesi tällaisen nukleotidin sisällyttämisen jälkeen keskeytyy. Näitä nukleotideja, joita kutsutaan dideoksinukleotideiksi, lisätään paljon vähemmän kuin tavallisesti, joten ketjun päättyminen tapahtuu vain satunnaisesti ja jokaisessa ketjussa eri paikoissa. Tuloksena on sekoitus eripituisia ketjuja, joista jokaisen päässä on sama nukleotidi. Siten ketjun pituus vastaa tutkitun sekvenssin nukleotidilukua, esimerkiksi jos meillä oli adenyylidideoksinukleotidi ja tuloksena saadut ketjut olivat 2, 7 ja 12 nukleotidiä pitkiä, niin adeniini oli toisessa, seitsemännessä ja kahdestoista paikassa geeni. Tuloksena oleva ketjuseos voidaan erottaa helposti koon mukaan elektroforeesilla, ja syntetisoidut ketjut voidaan tunnistaa radioaktiivisuudella röntgenfilmillä (katso kuva 10).

Osoittautuu kuvan alareunassa näkyvä kuva, nimeltään radioautograph. Liikkumalla sitä pitkin alhaalta ylös ja lukemalla kunkin vyöhykkeen sarakkeiden yläpuolella olevan kirjaimen, saamme nimikirjoituksen oikealla puolella olevassa kuvassa näkyvän nukleotidisekvenssin. Kävi ilmi, että synteesiä pysäyttävät dideoksinukleotidien lisäksi myös nukleotidit, joissa jonkinlainen kemiallinen ryhmä kuten fluoresoiva väriaine. Jos jokainen nukleotidi on leimattu omalla väriaineella, syntetisoidut ketjut erottamalla saadut vyöhykkeet hehkuvat eri valolla. Tämä mahdollistaa reaktion suorittamisen yhdessä koeputkessa samanaikaisesti kaikille nukleotideille ja erottamalla tuloksena olevat ketjut pituuden mukaan, tunnistaa nukleotidit värin perusteella (katso kuva 11).

Tällaiset menetelmät mahdollistivat yksittäisten geenien sekvenssien määrittämisen, mutta myös kokonaisten genomien lukemisen. Vielä nopeampia menetelmiä geenien nukleotidisekvenssien määrittämiseksi on nyt kehitetty. Jos ensimmäinen ihmisgenomi selvitettiin suuressa kansainvälisessä konsortiossa ensimmäisellä annetulla menetelmällä 12 vuodessa, toisella, toisella kolmessa vuodessa, nyt tämä voidaan tehdä kuukaudessa. Näin voit ennustaa henkilön alttiutta monille sairauksille ja ryhtyä toimenpiteisiin etukäteen niiden välttämiseksi.




