Alkueläinten erikoistunut organelli, jolla on eritystoiminto. Soluorganellit. Rakenne ja toiminnot. Missä eukaryooteissa seksuaalinen prosessi tapahtui ensimmäisen kerran?
Alkueläinperheeseen kuuluu noin 25 tuhatta yksisoluista eläinlajia, jotka elävät vedessä, maaperässä tai muiden eläinten ja ihmisten eliöissä. Koska solujen rakenteessa on morfologisia yhtäläisyyksiä monisoluisten organismien kanssa, alkueläimet eroavat niistä merkittävästi toiminnallisesti.
Jos monisoluisen eläimen solut suorittavat erityistoimintoja, niin alkueläimen solu on itsenäinen organismi, joka kykenee aineenvaihduntaan, ärtyneisyyteen, liikkumiseen ja lisääntymiseen.
Alkueläimet ovat organismeja solutasolla. Morfologisesti alkueläin vastaa solua, mutta fysiologisesti se on kokonainen itsenäinen organismi. Suurin osa niistä on kooltaan mikroskooppisesti pieniä (2-150 mikronia). Osa elävistä alkueläimistä saavuttaa kuitenkin 1 cm:n, ja useiden fossiilisten juurakoiden kuorien halkaisija on jopa 5-6 cm. Tunnettujen lajien kokonaismäärä on yli 25 tuhatta.
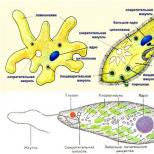
Alkueläinten rakenne on erittäin monipuolinen, mutta niillä kaikilla on solun organisaatiolle ja toiminnalle ominaisia piirteitä. Alkueläinten rakenteessa yleistä ovat kehon kaksi pääkomponenttia - sytoplasma ja ydin.

Sytaplasma
Sytoplasmaa rajoittaa ulkokalvo, joka säätelee aineiden virtausta soluun. Monissa alkueläimissä sitä monimutkaistavat lisärakenteet, jotka lisäävät ulkokerroksen paksuutta ja mekaanista lujuutta. Siten muodostuu muodostumia, kuten kalvoja ja kalvoja.
Alkueläinten sytoplasma on yleensä jaettu kahteen kerrokseen - ulompi on kevyempi ja tiheämpi - ektoplasma ja sisäinen, varustettu lukuisilla inkluusioilla, - endoplasma.
Yleiset soluelimet sijaitsevat sytoplasmassa. Lisäksi monien alkueläinten sytoplasmassa voi olla useita erityisiä organelleja. Erilaiset fibrillaariset muodostelmat ovat erityisen laajalle levinneitä - tuki- ja supistuvat kuidut, supistuvat tyhjiöt, ruoansulatusvakuolit jne.
Ydin
Alkueläimillä on tyypillinen soluydin, yksi tai useampi. Alkueläinten ytimellä on tyypillinen kaksikerroksinen ydinvaippa. Kromatiinimateriaali ja nukleolit jakautuvat ytimeen. Alkueläinten ytimille on ominaista poikkeuksellinen morfologinen monimuotoisuus koon, nukleolien lukumäärän, tumamehun jne. suhteen.
Alkueläinten elämäntoiminnan piirteet
Toisin kuin somaattiset solut, monisoluisille alkueläimille on ominaista elinkaaren läsnäolo. Se koostuu useista peräkkäisistä vaiheista, jotka toistuvat tietyllä kaavalla kunkin lajin olemassaolossa.
Useimmiten sykli alkaa tsygoottivaiheella, joka vastaa monisoluisten organismien hedelmöitettyä munaa. Tätä vaihetta seuraa yksi tai useampi toistuva suvuton lisääntyminen, joka suoritetaan solujakautumisen kautta. Sitten muodostuu sukupuolisoluja (sukusoluja), joiden pareittainen fuusio tuottaa jälleen tsygootin.
Monien alkueläinten tärkeä biologinen ominaisuus on kyky kirjoittelu. Tässä tapauksessa eläimet pyöristyvät, irrottavat tai vetäytyvät liikeorganellit, erittävät tiheän kuoren pinnalle ja joutuvat lepotilaan. Alkueläimet voivat sietää äkillisiä muutoksia ympäristössä säilyttäen samalla elinkelpoisuuden. Kun elämälle suotuisat olosuhteet palaavat, kystat avautuvat ja niistä poistuvat alkueläimet aktiivisina, liikkuvina yksilöinä.
Liikeorganellien rakenteen ja lisääntymisominaisuuksien perusteella alkueläintyyppi jaetaan 6 luokkaan. 4 pääluokkaa: Sarcodaceae, Flagellates, Sporozoans ja Ciliates.
Yksinkertaisimmat eläimet ovat yksisoluisia organismeja, ominaisuuksia, ravintoa, läsnäoloa vedessä ja ihmiskehossa
Yleiset luonteenpiirteet
Tai yksisoluiset organismit koostuvat nimensä mukaisesti yhdestä solusta. Alkueläinperheeseen kuuluu yli 28 000 lajia. Alkueläinten rakennetta voidaan verrata monisoluisten organismien solujen rakenteeseen. Molemmat perustuvat ytimeen ja sytoplasmaan, jossa on erilaisia organelleja (organelleja) ja sulkeumia. Emme kuitenkaan saa unohtaa, että mikä tahansa monisoluisen organismin solu on osa mitä tahansa kudosta tai elintä, jossa se suorittaa erityistehtävänsä. Kaikki monisoluisen organismin solut ovat erikoistuneita eivätkä kykene itsenäiseen olemassaoloon. Sitä vastoin yksinkertaisimmissa eläimissä yhdistyvät solun ja itsenäisen organismin toiminnot. (Fysiologisesti alkueläinsolu ei ole samanlainen kuin monisoluisten eläinten yksittäiset solut, vaan koko monisoluinen organismi.
Yksinkertaisin kaikki elävien organismien toiminnot ovat ominaisia: ravitsemus, aineenvaihdunta, erittyminen, ulkoisten ärsykkeiden havaitseminen ja reaktio niihin, liike, kasvu, lisääntyminen ja kuolema.
Alkueläimet Solun rakenne
Ydin ja sytoplasma, kuten on osoitettu, ovat minkä tahansa solun, mukaan lukien yksisoluiset eläimet, tärkeimmät rakenteelliset ja toiminnalliset komponentit. Jälkimmäisen runko sisältää organelleja, luusto- ja supistumiselementtejä sekä erilaisia sulkeumia. Se on aina peitetty solukalvolla, enemmän tai vähemmän ohuella, mutta selvästi näkyvällä elektronimikroskoopilla. Alkueläinten sytoplasma on nestemäistä, mutta sen viskositeetti vaihtelee eri lajien välillä ja vaihtelee eläimen kunnon ja ympäristön (sen lämpötilan ja kemiallisen koostumuksen) mukaan. Useimmissa lajeissa sytoplasma on läpinäkyvä tai maidonvalkoinen, mutta joissain se on väriltään sinistä tai vihertävää (Stentor, Fabrea saliva). Alkueläinten ytimen ja sytoplasman kemiallista koostumusta ei ole täysin tutkittu, mikä johtuu pääasiassa näiden eläinten pienestä koosta. Tiedetään, että sytoplasman ja ytimen perusta, kuten kaikissa eläimissä, koostuu proteiineista. Nukleiinihapot ovat läheistä sukua proteiineille, ne muodostavat nukleoproteiineja, joiden rooli kaikkien organismien elämässä on erittäin suuri. DNA (deoksiribonukleiinihappo) on osa alkueläinytimen kromosomeja ja varmistaa perinnöllisen tiedon siirtymisen sukupolvelta toiselle. RNA:ta (ribonukleiinihappoa) löytyy alkueläimistä sekä tumassa että sytoplasmassa. Se toteuttaa DNA:han koodattujen yksisoluisten organismien perinnöllisiä ominaisuuksia, koska sillä on johtava rooli proteiinien synteesissä.
Sytoplasman erittäin tärkeät kemialliset komponentit - rasvamaiset aineet lipidit - osallistuvat aineenvaihduntaan. Jotkut niistä sisältävät fosforia (fosfatideja), monet liittyvät proteiineihin ja muodostavat lipoproteiinikomplekseja. Sytoplasma sisältää myös vararavinteita sulkeumien - pisaroiden tai rakeiden - muodossa. Näitä ovat hiilihydraatit (glykogeeni, paramyyli), rasvat ja lipidit. Ne toimivat alkueläinkehon energiavarastona.
Orgaanisten aineiden lisäksi sytoplasmassa on runsaasti vettä ja mineraalisuoloja (kationit: K+, Ca2+, Mg2+, Na+, Fe3+ ja anionit: Cl~, P043“, N03“). Alkueläinten sytoplasmasta löytyy monia aineenvaihduntaan osallistuvia entsyymejä: proteaaseja, jotka varmistavat proteiinien hajoamisen; hiilihydraasit, jotka hajottavat polysakkarideja; lipaasit, jotka edistävät rasvansulatusta; suuri määrä entsyymejä, jotka säätelevät kaasunvaihtoa, nimittäin alkaliset ja happamat fosfataasit, oksidaasit, peroksidaasit ja sytokromioksidaasit.
Aiemmat käsitykset alkueläinten sytoplasman fibrillaarisesta, rakeisesta tai vaahto-solurakenteesta perustuivat tutkimuksiin kiinnitetyistä ja värjäytyneistä valmisteista. Uudet menetelmät alkueläinten tutkimiseksi (pimeässä kentässä, polarisoidussa valossa, intravitaalista värjäystä ja elektronimikroskopiaa käyttäen) ovat mahdollistaneet sen, että alkueläinten sytoplasma on monimutkainen dynaaminen hydrofiilisten kolloidien (pääasiassa proteiinikompleksien) järjestelmä, jolla on nestemäinen tai puolinestemäinen koostumus. Ultramikroskooppisessa tutkimuksessa pimeässä kentässä alkueläinten sytoplasma näyttää optisesti tyhjältä, vain soluorganellit ja niiden sulkeumat ovat näkyvissä.
Sytoplasmisten proteiinien kolloidinen tila varmistaa sen rakenteen vaihtelevuuden. Sytoplasmassa proteiinien aggregaattitilassa tapahtuu jatkuvasti muutoksia: ne siirtyvät nestemäisestä tilasta (sol) kiinteämpään, hyytelömäisempään tilaan (geeli). Nämä prosessit liittyvät tiheämmän ektoplasman kerroksen vapautumiseen, kuoren muodostumiseen - pellikeleihin ja monien alkueläinten ameboidiliikkeeseen.
Alkueläinten ytimet, kuten monisoluisten solujen ytimet, koostuvat kromatiinimateriaalista, tuman mehusta ja sisältävät tumasoluja ja tumakalvon. Useimmat alkueläimet sisältävät vain yhden ytimen, mutta on myös monitumaisia muotoja. Tässä tapauksessa ytimet voivat olla samoja (Pelomyxa-suvun monitumaiset amebat, moniytimiset siimat Polymastigida, Opalinida) tai eroavat muodoltaan ja toiminnaltaan. Jälkimmäisessä tapauksessa he puhuvat ydindifferentiaatiosta tai ydindualismista. Siten koko ripsien luokalle ja joillekin foraminifereille on ominaista ydindualismi. eli ytimet, jotka ovat muodoltaan ja toiminnaltaan epätasa-arvoisia.
Tämäntyyppiset alkueläimet, kuten muutkin organismit, noudattavat kromosomien lukumäärän pysyvyyden lakia. Niiden lukumäärä voi olla yksittäinen tai haploidinen (useimmat siima- ja itiöeläimet) tai kaksois- tai diploidi (ripset, opaalit ja ilmeisesti sarkodiat). Kromosomien lukumäärä eri alkueläinlajeissa vaihtelee suuresti: 2-4:stä 100-125:een (haploidisessa sarjassa). Lisäksi havaitaan ytimiä, joissa kromosomisarjojen lukumäärä on moninkertainen. Niitä kutsutaan polyploideiksi. Todettiin, että ripsien suuret ytimet tai makroytimet ja joidenkin radiolaarien ytimet ovat polyploideja. On hyvin todennäköistä, että myös Amoeba proteuksen ydin on polyploidinen; tämän lajin kromosomien lukumäärä on 500.
Lisääntyminen Ydinjako
Pääasiallinen tuman jakautumisen tyyppi sekä alkueläimissä että monisoluisissa organismeissa on mitoosi tai karyokineesi. Mitoosin aikana kromosomimateriaalin oikea, tasainen jakautuminen tapahtuu jakautuvien solujen tumien välillä. Tämä varmistetaan kunkin kromosomin pituussuunnassa jakautumisella kahdeksi tytärkromosomiin mitoosin metafaasissa, jolloin molemmat tytärkromosomit menevät jakautuvan solun eri napoihin.
 Monocystis magnan gregarin ytimen mitoottinen jakautuminen:
Monocystis magnan gregarin ytimen mitoottinen jakautuminen: 1, 2 - profaasi; 3 - siirtyminen metafaasiin; 4, 5 - metafaasi; 6 - varhainen anafaasi; 7, 8 - myöhässä
anafaasi; 9, 10 - telofaasi.
Monocystis magna gregarina -tuman jakautuessa voidaan havaita kaikki monisoluisille organismeille tyypilliset mitoottiset hahmot. Profaasissa ytimessä näkyy lankamaisia kromosomeja, joista osa liittyy ytimeen (kuvat 1, 1, 2). Sytoplasmassa voidaan erottaa kaksi sentrosomia, joiden keskellä on sentrioleja, joiden tähtisäteet eroavat säteittäisesti. Centrosomit lähestyvät ydintä, liittyvät sen kuoreen ja siirtyvät ytimen vastakkaisille napoille. Ydinvaippa liukenee ja muodostuu akromatiinikara (Kuva 1, 2-4). Kromosomien spiralisoituminen tapahtuu, minkä seurauksena ne lyhentyvät suuresti ja kerääntyvät ytimen keskelle, ydin liukenee. Metafaasissa kromosomit siirtyvät ekvatoriaaliselle tasolle. Jokainen kromosomi koostuu kahdesta kromatidista, jotka sijaitsevat yhdensuuntaisesti toistensa kanssa ja joita pitää yhdessä yksi sentromeeri. Tähtihahmo kunkin senrosomin ympäriltä katoaa ja sentriolit jakautuvat kahtia (kuvat 1, 4, 5). Anafaasissa kunkin kromosomin sentromeerit jakautuvat kahtia ja niiden kromatidit alkavat hajota kohti karan napoja. Alkueläimille on ominaista, että sentromeereihin kiinnittyneet vetokarafilamentit ovat erotettavissa vain joissakin lajeissa. Koko kara on venynyt ja sen kierteet, jotka kulkevat jatkuvasti napasta napaan, pidentyvät. Kromosomeiksi muuttuneiden kromatidien erottaminen varmistetaan kahdella mekanismilla: niiden vetäytyminen irti vetokaran kierteiden supistumisen vaikutuksesta ja jatkuvien karan kierteiden venyminen. Jälkimmäinen johtaa solunapojen irtoamiseen toisistaan (kuvat 1, 6, 7) Telofaasissa prosessi etenee päinvastaisessa järjestyksessä: jokaisessa navassa joukko kromosomeja puetaan ydinvaippaan. kromosomit hajoavat ja ohenevat, ja muodostuu uudelleen nukleoleja. Kara katoaa ja jakautuneiden sentriolien ympärille muodostuu kaksi itsenäistä sentrosomia tähtisäteineen. Jokaisessa tytärsolussa on kaksi sentrosomia - seuraavan mitoottisen jakautumisen tulevat keskukset (kuva 1, 9, 10). Tuman jakautumisen jälkeen sytoplasma yleensä jakautuu. Kuitenkin joissakin alkueläimissä, mukaan lukien Monocystiksessa, tapahtuu sarja peräkkäisiä tuman jakautumista, joiden seurauksena elinkaareen syntyy tilapäisesti moniytimiä. sytoplasman osa eristetään jokaisen ytimen ympäriltä ja useita pieniä soluja muodostuu samanaikaisesti.
Yllä kuvatusta mitoosiprosessista on erilaisia poikkeamia: ydinvaippa voi säilyä koko mitoottisen jakautumisen ajan, ydinvaipan alle voi muodostua akromatiinikara, ja joissain muodoissa sentrioleja ei muodostu. Merkittävimmät poikkeamat ovat joissakin euglenidae-lajeissa: niiltä puuttuu tyypillinen metafaasi ja kara kulkee ytimen ulkopuolelle. Metafaasissa kahdesta kromatidista koostuvat kromosomit sijaitsevat ytimen akselia pitkin, ekvatoriaalista levyä ei muodostu, ydinkalvo ja ydin säilyvät, jälkimmäinen jaetaan puoliksi ja siirtyy tytärytimiin. Alkueläinten ja monisoluisten organismien mitoosissa olevien kromosomien käyttäytymisessä ei ole perustavanlaatuisia eroja.
Ennen uusien tutkimusmenetelmien käyttöä monien alkueläinten ydinjakautumista kuvattiin amitoosiksi eli suoraksi jakautumiseksi. Todellinen amitoosi ymmärretään nykyään ytimien jakautumiseksi ilman kromatidien (kromosomien) asianmukaista erottamista tytärytimiin. Tämän seurauksena muodostuu ytimiä, joissa on epätäydellisiä kromosomisarjoja. Ne eivät pysty jatkamaan normaalia mitoottista jakautumista. Tällaisia ydinjakaumia on vaikea odottaa normaalisti yksinkertaisimmissa organismeissa. Amitoosi havaitaan valinnaisesti enemmän tai vähemmän patologisena prosessina.
Alkueläinten runko on melko monimutkainen. Yhden solun sisällä tapahtuu sen yksittäisten osien erilaistumista, jotka suorittavat erilaisia toimintoja. Siten, analogisesti monisoluisten eläinten elinten kanssa, näitä alkueläinten osia kutsuttiin organelleiksi tai organelleiksi. Siellä on liikkeen, ravinnon, valon ja muiden ärsykkeiden havaitsemisen organelleja, erittäviä organelleja jne.
Liike
Alkueläinten liikeelimet ovat pseudopodia tai pseudopodit, siimot ja värekärpit. Pseudopodia muodostuu suurimmaksi osaksi liikehetkellä ja ne voivat kadota heti, kun alkueläin lakkaa liikkumasta. Pseudopodia ovat tilapäisiä plasmamaisia alkueläinten kehon kasvuja, joilla ei ole pysyvää muotoa. Niiden kuorta edustaa erittäin ohut (70-100 A) ja elastinen solukalvo. Pseudopodiat ovat tyypillisiä sarkoodeille, joillekin siimoille ja itiöeläimille.
Flagella ja värekarvat ovat sytoplasman ulkokerroksen pysyviä kasvaimia, jotka kykenevät rytmisiin liikkeisiin. Näiden organellien ultrahienoa rakennetta tutkittiin elektronimikroskoopilla. Todettiin, että ne on rakennettu pitkälti samalla tavalla. Siiman tai ciliumin vapaa osa ulottuu solun pinnasta.
Sisäosa on upotettu ektoplasmaan ja sitä kutsutaan tyvikappaleeksi tai blefaroplastiksi. Siiman tai ciliumin ultraohuilla osilla voidaan erottaa 11 pitkittäistä fibrilliä, joista 2 sijaitsee keskellä ja 9 reunalla (kuva 2). Joidenkin lajien keskusfibrilleissä on kierteisiä juovia. Jokainen perifeerinen fibrilli koostuu kahdesta yhdistetystä putkesta tai subfbrillistä. Perifeeriset fibrillit kulkeutuvat perusrunkoon, mutta keskeiset fibrillit eivät saavuta sitä. Siimakalvo siirtyy alkueläinrungon kalvoon.
Huolimatta värien ja flagellan rakenteen samankaltaisuudesta, niiden liike on erilainen. Jos flagellat tekevät monimutkaisia ruuviliikkeitä, värekkojen työtä voidaan verrata helpoimmin airojen liikkeisiin.
Joidenkin alkueläinten sytoplasmassa on perusrungon lisäksi parabasaalirunko. Perusrunko on koko tuki- ja liikuntaelimistön perusta; lisäksi se säätelee alkueläinten mitoottista jakautumisprosessia. Parabasaalirunko osallistuu alkueläinten aineenvaihduntaan; toisinaan se katoaa ja voi sitten ilmestyä uudelleen.
Tuntoelimet
Alkueläimet pystyvät määrittämään valon intensiteetin (valaistuksen) käyttämällä valoherkkää organellia - ocellusta. Merisiimalajin Chromulina psammobian silmän ultraohuen rakenteen tutkimus osoitti, että se sisältää muunnetun siimasolun upotettuna sytoplasmaan.
Erilaisten ravintotyyppien yhteydessä, joita käsitellään tarkemmin myöhemmin, alkueläimillä on hyvin laaja valikoima ruoansulatuselimet: yksinkertaisista ruuansulatusvakuoleista tai rakkuloista sellaisiin erikoistuneisiin muodostelmiin kuten solusuu, suusuppilo, nielu, jauhe.
Eritysjärjestelmä
Useimmille alkueläimille on ominaista kyky kestää epäsuotuisia ympäristöolosuhteita (tilapäisten säiliöiden kuivuminen, lämpö, kylmä jne.) kystien muodossa. Alkueläin vapauttaa merkittävän määrän vettä valmistautuessaan entisaatioon, mikä johtaa sytoplasman tiheyden lisääntymiseen. Ruokahiukkasten jäännökset heitetään ulos, värekarvot ja flagellat katoavat ja pseudopodia vedetään sisään. Kokonaisaineenvaihdunta heikkenee, muodostuu suojaava kuori, joka koostuu usein kahdesta kerroksesta. Kystien muodostumista monissa muodoissa edeltää vararavinteiden kertyminen sytoplasmaan.
Alkueläimet eivät menetä elinkelpoisuutta kystissä kovin pitkään. Kokeissa nämä ajanjaksot ylittivät 5 vuotta Oicomonas-suvun (Protomonadida), 8 vuotta Haematococcus pluvialis -suvun osalta ja Peridinium cinctumin kystien enimmäiseloonjäämisaika ylitti 16 vuotta.
Tuuli kuljettaa kystien muodossa alkueläimiä pitkiä matkoja, mikä selittää alkueläineläimistön homogeenisuuden kaikkialla maapallolla. Siten kystillä ei ole vain suojaavaa tehtävää, vaan ne toimivat myös alkueläinten pääasiallisena leviämiskeinona.
Soluelimet, jotka tunnetaan myös organelleina, ovat itse solun erikoistuneita rakenteita, jotka vastaavat useista tärkeistä ja elintärkeistä toiminnoista. Miksi sitten "organellit"? On vain niin, että täällä näitä solukomponentteja verrataan monisoluisen organismin elimiin.
Mitkä organellit muodostavat solun?
Myös joskus organellit tarkoittavat vain solun pysyviä rakenteita, jotka sijaitsevat siinä. Samasta syystä solun ydintä ja sen ytimeä ei kutsuta organelleiksi, kuten myös värekarvot ja flagellat eivät ole organelleja. Mutta solun muodostaviin organelleihin kuuluvat: kompleksi, endoplasminen verkkokalvo, ribosomit, mikrotubulukset, mikrofilamentit, lysosomit. Itse asiassa nämä ovat solun tärkeimmät organellit.
Jos puhumme eläinsoluista, niiden organelleihin kuuluu myös sentrioleja ja mikrofibrillejä. Mutta kasvisolun organellien lukumäärä sisältää edelleen vain kasveille ominaisia plastideja. Yleensä solujen organellien koostumus voi vaihdella merkittävästi riippuen itse solutyypistä.

Piirustus solun rakenteesta, mukaan lukien sen organellit.
Kaksoiskalvosoluorganellit
Myös biologiassa on sellainen ilmiö kuin kaksoiskalvosoluorganellit, joita ovat mitokondriot ja plastidit. Alla kuvailemme niiden luontaisia toimintoja sekä kaikkia muita tärkeimpiä organelleja.
Soluelinten toiminnot
Kuvailkaamme nyt lyhyesti eläinsolujen organellien päätoimintoja. Niin:
- Plasmakalvo on ohut kalvo solun ympärillä, joka koostuu lipideistä ja proteiineista. Erittäin tärkeä organelli, joka kuljettaa vettä, mineraaleja ja orgaanisia aineita soluun, poistaa haitallisia kuona-aineita ja suojaa solua.
- Sytoplasma on solun sisäinen puolinesteympäristö. Tarjoaa yhteyden ytimen ja organellien välillä.
- Endoplasminen verkkokalvo on myös kanavien verkosto sytoplasmassa. Osallistuu aktiivisesti proteiinien, hiilihydraattien ja lipidien synteesiin ja ravintoaineiden kuljettamiseen.
- Mitokondriot ovat organelleja, joissa orgaaniset aineet hapetetaan ja ATP-molekyylejä syntetisoidaan entsyymien osallistuessa. Pohjimmiltaan mitokondriot ovat soluorganellit, jotka syntetisoivat energiaa.
- Plastidit (kloroplastit, leukoplastit, kromoplastit) - kuten edellä mainittiin, niitä löytyy yksinomaan kasvisoluista; yleensä niiden läsnäolo on kasviorganismin pääominaisuus. Niillä on erittäin tärkeä tehtävä, esimerkiksi kloroplastit, jotka sisältävät vihreää pigmenttiä klorofylliä, ovat vastuussa ilmiöstä kasveissa.
- Golgi-kompleksi on sytoplasmasta kalvolla rajattu onteloiden järjestelmä. Suorita rasvojen ja hiilihydraattien synteesi kalvolla.
- Lysosomit ovat soluja, jotka on erotettu sytoplasmasta kalvolla. Niiden sisältämät erityiset entsyymit nopeuttavat monimutkaisten molekyylien hajoamista. Lysosomi on myös organelli, joka varmistaa proteiinien kokoamisen soluissa.
- - solunesteellä täytetyt sytoplasman ontelot, vararavinteiden kertymispaikka; ne säätelevät solun vesipitoisuutta.
Yleensä kaikki organellit ovat tärkeitä, koska ne säätelevät solun elämää.
Perussoluorganellit, video
Ja lopuksi temaattinen video soluorganelleista.
Jokainen elävä organismi koostuu soluista, joista monet pystyvät liikkumaan. Tässä artikkelissa puhumme liikeorganelleista, niiden rakenteesta ja toiminnoista.
Yksisoluisten organismien liikeorganellit
Nykyaikaisessa biologiassa solut jaetaan prokaryooteihin ja eukaryooteihin. Ensimmäiset sisältävät edustajia yksinkertaisimmista organismeista, jotka sisältävät yhden DNA-juosteen ja joilla ei ole ydintä (sinilevät, virukset).
Eukaryooteilla on ydin ja ne koostuvat useista organelleista, joista yksi on liikeorganellit.
Yksisoluisten organismien liikeorganelleja ovat värekarvot, flagellat, lankamaiset muodostelmat - myofibrillit, pseudopods. Niiden avulla solu voi liikkua vapaasti.

Riisi. 1. Liikuntaelinten lajikkeet.
Liikeorganelleja löytyy myös monisoluisista organismeista. Esimerkiksi ihmisillä keuhkoputkien epiteeli on peitetty monilla väreillä, jotka liikkuvat tiukasti samassa järjestyksessä. Tässä tapauksessa muodostuu niin kutsuttu "aalto", joka voi suojata hengitysteitä pölyltä ja vierailta hiukkasilta. Siittiöissä (urospuolisen kehon erikoistuneet solut, jotka palvelevat lisääntymistä) on myös siimoja.
TOP 4 artikkeliajotka lukevat tämän mukana
Motorinen toiminto voidaan suorittaa myös mikrokuitujen (myoneemien) supistumisen vuoksi, jotka sijaitsevat sytoplasmassa ihon alla.
Liikeelinten rakenne ja toiminnot
Liikeorganellit ovat kalvon kasvamia, joiden halkaisija on 0,25 µm. Rakenteeltaan siimat ovat paljon pidempiä kuin värekarvot.
Joidenkin nisäkkäiden siittiöiden siiman pituus voi olla 100 mikronia, kun taas värekkojen koko on jopa 15 mikronia.
Tällaisista eroista huolimatta näiden organellien sisäinen rakenne on täysin sama. Ne muodostuvat mikrotubuluksista, jotka ovat rakenteeltaan samanlaisia kuin solukeskuksen sentriolit.
Moottoriliikkeet muodostuvat mikrotubulusten liukumisesta keskenään, minkä seurauksena ne taipuvat. Näiden organellien pohjassa on perusrunko, joka kiinnittää ne solun sytoplasmaan. Liikuntaelinten toiminnan varmistamiseksi solu kuluttaa ATP-energiaa.

Riisi. 2. Siiman rakenne.
Jotkut solut (amebat, leukosyytit) liikkuvat pseudopodian, toisin sanoen pseudopodian, vuoksi. Toisin kuin siimot ja värekärpät, pseudopodia ovat väliaikaisia rakenteita. Ne voivat kadota ja esiintyä eri paikoissa sytoplasmassa. Niiden toimintoihin kuuluu liikkuminen sekä ruoan ja muiden hiukkasten talteenotto.
Flagella koostuu filamentista, koukusta ja tyvirungosta. Näiden organellien lukumäärän ja sijainnin mukaan bakteerien pinnalla ne on jaettu:
- Monotrichit(yksi siima);
- Amphitrichy(yksi siima eri navoissa);
- Lophotrichs(joukko muodostelmia yhdessä tai molemmissa navoissa);
- Peritrichous(monet siimat sijaitsevat koko solun pinnalla).

Riisi. 3. Siimalajikkeet.
Liikeorganellien suorittamien toimintojen joukossa ovat:
- liikkeen tarjoaminen yksisoluiselle organismille;
- lihasten kyky supistua;
- hengitysteiden suojaava reaktio vierailta hiukkasilta;
- nesteen eteneminen.
Flagellateilla on suuri rooli ympäristön aineiden kierrossa, ja monet niistä ovat hyviä indikaattoreita vesistöjen saastumisesta.
Mitä olemme oppineet?
Yksi solun peruselementeistä on liikeorganellit. Näitä ovat siimat ja värekarvot, jotka muodostuvat mikrotubulusten avulla. Niiden toimintoihin kuuluu liikkeen tarjoaminen yksisoluiselle organismille ja nesteiden edistäminen monisoluisen organismin sisällä.
Testi aiheesta
Raportin arviointi
Keskiarvoluokitus: 4.7. Saatujen arvioiden kokonaismäärä: 175.
Organelli on pysyvä muodostus solussa, joka suorittaa tiettyjä toimintoja. Niitä kutsutaan myös organelleiksi. Organelli on se, mikä sallii solun elää. Aivan kuten eläimet ja ihmiset koostuvat elimistä, niin jokainen solu koostuu organelleista. Ne ovat monipuolisia ja suorittavat kaikki toiminnot, jotka varmistavat solun elämän: aineenvaihdunta, varastointi ja jakautuminen.
Millaisia organelleja on olemassa?
Organelli on monimutkainen rakenne. Joillakin heistä voi jopa olla oma DNA ja RNA. Kaikki solut sisältävät mitokondrioita, ribosomeja, lysosomeja, solukeskuksen, Golgi-laitteiston (kompleksin) ja endoplasmisen retikulumin (verkkokalvon). Kasveilla on myös erityisiä soluorganelleja: vakuoleja ja plastideja. Jotkut myös luokittelevat mikrotubulukset ja mikrofilamentit organelleiksi.
Organelli on ribosomi, vakuoli, solukeskus ja monet muut. Katsotaanpa tarkemmin organellien rakennetta ja toimintoja.
Mitokondriot
Nämä organellit tarjoavat solulle energiaa - ne ovat vastuussa niistä. Niitä löytyy kasveista, eläimistä ja sienistä. Näillä soluorganelleilla on kaksi kalvoa: ulkoinen ja sisäinen, joiden välissä on kalvojen välinen tila. Sitä, mikä on kuorien sisällä, kutsutaan matriisiksi. Se sisältää erilaisia entsyymejä - aineita, jotka ovat välttämättömiä kemiallisten reaktioiden nopeuttamiseksi. Sisäkalvossa on poimuja, joita kutsutaan cristaeiksi. Juuri heillä tapahtuu soluhengitysprosessi. Lisäksi mitokondriomatriisi sisältää mitokondriaalista DNA:ta (mDNA) ja mRNA:ta sekä ribosomeja, jotka ovat melkein samanlaisia kuin mitokondrioiden omat ribosomit.
Ribosomi
Tämä organelli on vastuussa translaatioprosessista, jossa proteiini syntetisoidaan yksittäisistä aminohapoista. Ribosomiorganellin rakenne on yksinkertaisempi kuin mitokondrioiden - siinä ei ole kalvoja. Tämä organelli koostuu kahdesta osasta (alayksiköistä) - pienestä ja suuresta. Kun ribosomi on inaktiivinen, ne erotetaan, ja kun se alkaa syntetisoida proteiineja, ne yhdistyvät. Useita ribosomeja voi myös yhdistyä, jos niiden syntetisoima polypeptidiketju on hyvin pitkä. Tätä rakennetta kutsutaan "polyribosomiksi".

Lysosomit
Tämän tyyppisten organellien toiminnot rajoittuvat solujen ruoansulatukseen. Lysosomeilla on yksi kalvo, jonka sisällä on entsyymejä, jotka katalysoivat kemiallisia reaktioita. Joskus nämä organellit eivät vain hajoa, vaan myös sulattavat kokonaisia organelleja. Tämä voi tapahtua solun pitkäaikaisen nälänhädän aikana ja antaa sen elää jonkin aikaa. Vaikka ravinteet eivät vieläkään ala virrata, solu kuolee.

ja toimintoja
Tämä organelli koostuu kahdesta osasta - sentrioleista. Nämä ovat sylinterin muotoisia muodostelmia, jotka koostuvat mikrotubuluksista. Solukeskus on erittäin tärkeä organelli. Se on mukana karan muodostumisprosessissa. Lisäksi se on mikrotubulusten organisoinnin keskus.
Golgin laite
Se on kompleksi levymäisiä kalvopusseja, joita kutsutaan cisternaiksi. Tämän organellin toimintoihin kuuluu tiettyjen aineiden lajittelu, varastointi ja muuntaminen. Hiilihydraatit, jotka ovat osa glykokaliksia, syntetisoidaan täällä pääasiassa.

Endoplasmisen retikulumin rakenne ja toiminnot
Se on putkien ja taskujen verkosto, jota ympäröi yksi kalvo. Endoplasmista retikulumia on kahta tyyppiä: sileä ja karkea. Ribosomit sijaitsevat jälkimmäisen pinnalla. Sileä ja karkea verkkokalvo suorittaa erilaisia toimintoja. Ensimmäinen on vastuussa hormonien synteesistä, hiilihydraattien varastoinnista ja muuntamisesta. Lisäksi siihen muodostuu vakuolien alkioita, kasvisoluille tyypillisiä organelleja. Karkea endoplasminen verkkokalvo sisältää pinnallaan ribosomeja, jotka tuottavat aminohappojen polypeptidiketjun. Sitten se tulee endoplasmiseen retikulumiin, ja täällä muodostuu tietty proteiinin sekundaarinen, tertiäärinen ja kvaternäärinen rakenne (ketju kierretään oikealla tavalla).

Vacuoles
Nämä ovat organelleja, joissa on yksi kalvo. Niihin kertyy solumehua. Vakuoli on välttämätön turgorin ylläpitämiseksi. Se osallistuu myös osmoosiprosessiin. Lisäksi niitä löytyy pääasiassa vesistöissä elävistä yksisoluisista organismeista, ja ne toimivat pumpuina, jotka pumppaavat pois ylimääräistä nestettä solusta.
Plastidit: lajikkeet, rakenne ja toiminnot
Nämä ovat myös organelleja, joita on kolmea tyyppiä: leukoplastit, kromoplastit ja kloroplastit. Ensimmäiset varastoivat vararavinteita, pääasiassa tärkkelystä. Kromoplastit sisältävät erilaisia pigmenttejä. Niiden ansiosta kasvien terälehdet ovat monivärisiä. Keho tarvitsee tätä ensisijaisesti pölyttävien hyönteisten houkuttelemiseksi.
Kloroplastit ovat tärkeimpiä plastideja. Suurin määrä niitä löytyy kasvien lehdistä ja varresta. He ovat vastuussa fotosynteesistä - kemiallisten reaktioiden ketjusta, jonka aikana keho tuottaa orgaanisia aineita epäorgaanisista aineista. Näillä organelleilla on kaksi kalvoa. Kloroplastien matriisia kutsutaan "stroomaksi". Se sisältää plastidi-DNA:ta, RNA:ta, entsyymejä ja tärkkelyssulkeumia. Kloroplastit sisältävät tylakoideja, kolikon muotoisia kalvomuodostelmia. Niiden sisällä tapahtuu fotosynteesi. Se sisältää myös klorofylliä, joka toimii kemiallisten reaktioiden katalysaattorina. Kloroplastien tylakoidit yhdistetään pinoiksi - granaksi. Organellit sisältävät myös lamelleja, jotka yhdistävät yksittäisiä tylakoideja ja tarjoavat yhteyden niiden välillä.
Liikeorganellit
Ne ovat ominaisia pääasiassa yksisoluisille organismeille. Näitä ovat siimat ja värekarvet. Ensimmäisiä esiintyy euglenoissa, trypanosomeissa ja klamydomonasissa. Flagellaa on myös eläinten siittiöissä. Särmäsoluilla ja muilla yksisoluisilla organismeilla on värpäreitä.
Mikrotubulukset
Ne varmistavat aineiden kuljetuksen sekä solun jatkuvan muodon. Jotkut tutkijat eivät luokittele mikrotubuluksia organelleiksi.